Das reale Untersuchungsgebiet im südlichen Hochschwarzwald umfasst eine von fragmentiertem Bergmischwald dominierte Fläche von ca. 500 km² mit einer zentralen und für den Artenschutz relevanten Zielfläche von 3 000 ha, die aus dem tatsächlichen
Auerhuhnvorkommen im Gebiet (Braunisch, Suchant 2006) abgeleitet wurde (Abb. 2a). In der Auswertung wurden reale Jagdreviergrenzen verwendet, um die unregelmäßige Verzahnung von
Besitzverhältnissen auf der Fläche abzubilden. Die durchschnittliche Größe der Jagdreviere war sehr unterschiedlich (mit Standardabweichung von ca. 400 ha um
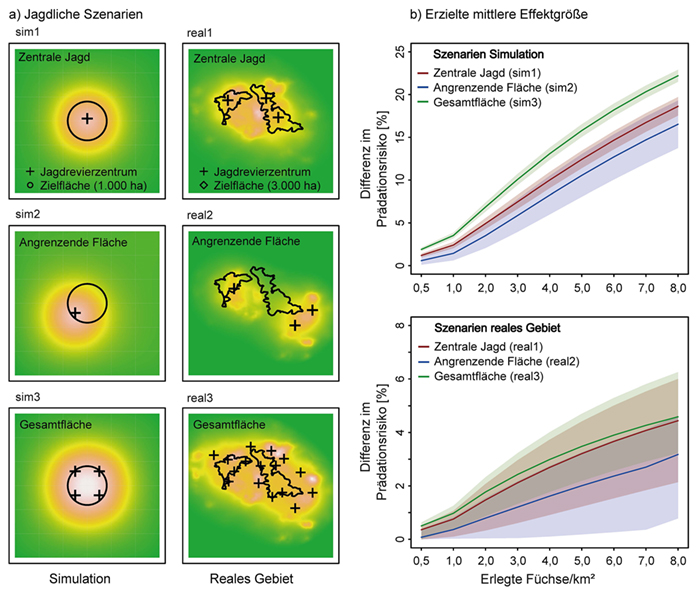
Abb. 2: a) Visualisierung der drei jagdlichen Szenarien für die Bewertung der Wirksamkeit von Fuchsbejagung auf Jagdrevierebene in Simulation (sim1, sim2, sim3) und realem Untersuchungsgebiet (real1, real2, real3). Farbliche Intensitäten zeigen die
erzeugte Effektstärke nach topographischem Farbschema (weiß = höchste Effektstärke; grün = keine Veränderung im Prädationsrisiko; separat für die Szenarien im realen Gebiet bzw. der Simulation). Für den räumlichen Bezug der
Bejagungsschwerpunkte siehe Methodenteil (Abschnitt 2). b) Erzielte mittlere Effektgröße (Differenz im Prädationsrisiko) über die Zielfläche mit 95 %-Quantilbereich in Abhängigkeit von zunehmender jagdlicher
Intensität in den beteiligten Revieren in allen Szenarien (siehe a). Eine koordinierte Jagd auf der Gesamtfläche führt jeweils zum größten mittleren Effekt bei geringster Streuung der Effektgröße über die Zielfläche (vgl. Jagd auf der Gesamtfläche in Simulation und
realem Gebiet).
Fig. 2: a) Spatial model predictions of predator removal effectiveness (quantified as the change in predation risk at a site) following three scenarios for predator removal and for a simulated (“Simulation”; sim1, sim2, sim3) and real study area
(“Reales Gebiet”; real1, real2, real3). Effect size corresponds to colour intensity along a topographic colour scheme (white = highest effect size; green = no difference in predation risk; separately for real and simulated study area). Crosses mark
focal sites for removal effort in the models. b) Mean achieved effect size (difference in predation risk) across the whole target area (see lines in a) with 95 % quantile range, as a function of increasing removal intensity at the focal sites. A
coordinated removal of foxes in both the target site and in adjacent concessions had the largest mean effect and the smallest variance both in the simulation and the real study area (compare sim3 and real3).
Simuliertes Untersuchungsgebiet
Das simulierte Untersuchungsgebiet entsprach im Verhältnis von Zielfläche zu Gesamtgröße dem realen Untersuchungsgebiet, jedoch mit deutlich reduzierter Komplexität. Es bestand aus 36 quadratischen Jagdrevieren mit einer Größe von jeweils 500 ha (gesamt
180 km²) mit einer zentralen Zielfläche von 1 000 ha in einer gleichförmigen Landschaft. Die Fuchsdichte im Gebiet wurde als konstant angenommen und auf den Mittelwert des Index der Fuchsdichte im realen Gebiet gesetzt (Kämmerle
et al. 2019b).
2.2 Zugrunde liegende Modelle
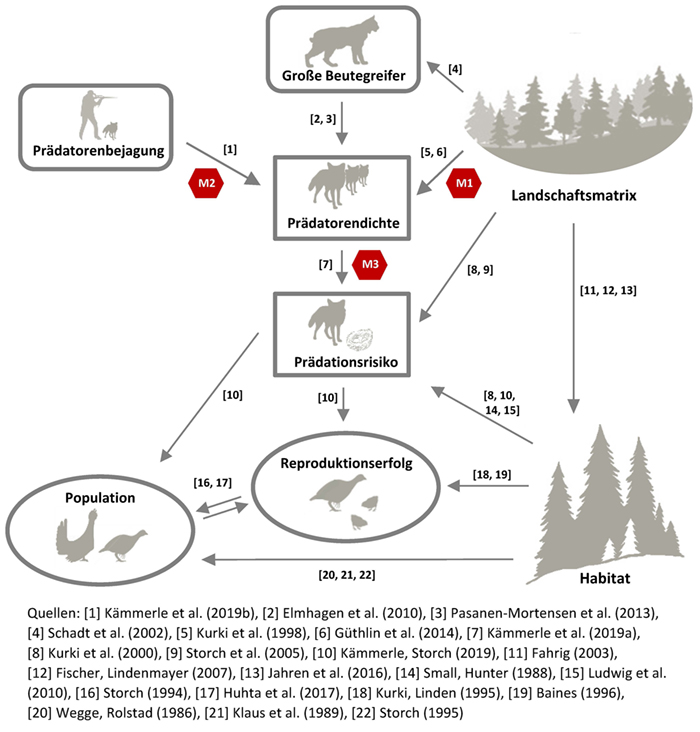
Modell M1: Fuchsdichte und Bejagungsintensität
Um Unterschiede in der Fuchsdichte innerhalb des realen Untersuchungsgebiets vorherzusagen, haben wir auf ein in diesem Gebiet entwickeltes Landschaftsmodell zurückgegriffen (Güthlin et al. 2013, 2014). Dieses Modell quantifiziert Unterschiede in der Fuchshäufigkeit basierend auf vier zentralen Einflussgrößen: Fragmentierungsgrad, Diversität der Landnutzung, Produktivität und Bodenqualität. Das Modell ist damit zwar räumlich, aber nicht
zeitlich explizit (d. h. keine Darstellung der jährlichen Dynamik in der Fuchsdichte). Das von uns genutzte Modell (Güthlin et al. 2014) sagt Unterschiede in einem relativen Häufigkeitsindex (RAI) vorher (Tab. 1) und basiert auf Daten aus Kamerafallen.
Unterschiede in der Bejagungsintensität und deren räumliche Effekte wurden mittels räumlicher Interpolation der standardisierten Jagdstrecken (d. h. erlegte Füchse pro km² Revierfläche) abgebildet. Es wurde eine flexible Oberfläche erzeugt, die durch die
Zentralpunkte aller Jagdreviere verläuft (im R-Paket „fields“; Nychka et al. 2017). Dieser Ansatz wurde gewählt, um die Anforderungen des Folgemodells zu erfüllen, das Unterschiede in der Fuchshäufigkeit als Folge der Jagd mittels
einer solchen Oberfläche quantifiziert (Kämmerle et al. 2019b).
Modell M2: Einfluss der Fuchsbejagung
Der Einfluss der Fuchsjagd auf die Fuchshäufigkeit wurde mittels eines Modells quantifiziert, das mithilfe auf Landschaftsebene erhobener Kamerafallendaten trainiert wurde (Kämmerle et al. 2019b). Dieses Modell kombiniert den
vorhergesagten relativen Index der Fuchshäufigkeit und die jagdliche Intensität (Interpolation) als unabhängige Variablen und sagt damit einen veränderten Index der Fuchshäufigkeit voraus.
Modell M3: Veränderung im Prädationsrisiko
Im letzten Schritt wurden Veränderungen im Prädationsrisiko als Folge jagdlicher Effekte auf die Fuchsdichte mittels eines Modells für die Wahrscheinlichkeit der Kunstgelegeprädation vorhergesagt (Kämmerle et al. 2019a). Dieses
Modell nutzte wiederum die in Modell M2 vorhergesagten Änderungen im relativen Index der Fuchshäufigkeit als Prädiktor basierend auf einem linearen Zusammenhang zwischen Fuchshäufigkeit und Kunstnestprädation (Kämmerle et al.
2019a).
Weitere Variablen (z. B. Plot-Charakteristika, Vegetationsbedeckung) wurden in dieser Auswertung konstant gehalten. Detaillierte Informationen zu den Modellen und deren Parametern sind den zitierten Primärveröffentlichungen zu entnehmen, die alle als
Open-Access-Publikationen online verfügbar sind.
2.3 Bejagungs-Szenarien
Es wurden drei jagdliche Strategien auf ihre Wirksamkeit untersucht, die denkbaren Ansätzen in der Praxis entsprechen:
● Szenario 1: eine Jagd zentral auf der Zielfläche,
● Szenario 2: eine intensive Bejagung in einzelnen Jagdrevieren angrenzend an die Zielfläche und
● Szenario 3: eine koordinierte Jagd auf der Gesamtfläche der Zielfläche inklusive aller angrenzenden Jagdreviere.
Die Modellkette wurde für jedes Szenario angewandt, jeweils für das reale Untersuchungsgebiet (Szenarien real1, real2, real3) sowie in vereinfachter Form für den Simulationsansatz (Szenarien sim1, sim2, sim3). Die räumliche Verortung der Szenarien auf der Fläche ist in Abb. 2 dargestellt.
Für jedes Szenario wurden jeweils neun Unterszenarien gerechnet, in denen die jagdliche Intensität auf den bejagten Flächen jeweils zwischen 0,5 und 8,0 erlegten Füchsen pro km² variierte (54 Kombinationen; Werte von ≥ 5 Füchsen/km² entsprachen
einer starken Bejagung im realen Gebiet). Die tatsächliche Fuchsdichte im realen Gebiet ist nicht bekannt, mittlere Streifgebietsgrößen von ≥ 200 ha (Kaphegyi 2002) weisen jedoch auf eine geschätzte Spanne von
2 – 5 Füchsen/km² hin (vgl. Šálek et al. 2015).
Die jagdliche Intensität auf Füchse wurde den Jagdrevieren, die als aktiv jagend gewertet wurden, entsprechend dem jeweiligen Szenario an deren Mittelpunkt zugewiesen (Kreuze in Abb. 2) und anschließend interpoliert
(alle unbeteiligten Reviere erhielten den Wert 0). Eine Ausnahme bilden sim1 und real1, in denen die Bejagung zentral auf der Zielfläche verortet wurde. Für die Szenarien 1 und 2 wurde jeweils ein Bejagungsschwerpunkt pro 1 000 ha Zielfläche
ausgewählt, in Szenario 3 wurden alle angrenzenden Reviere selektiert (d. h. der jagdliche Aufwand war höher).
Die räumlichen Effekte wurden jeweils bei einer jagdlichen Intensität von 6 erlegten Füchsen pro km² visualisiert (entsprechend einem sehr hohen Wert im realen Untersuchungsgebiet). Die Veränderung im Prädationsrisiko innerhalb der Zielfläche als Folge der Jagd wurde
schließlich mit dem vorhergesagten Prädationsrisiko bezogen auf die Fuchsdichte im Gebiet vor der Jagd verglichen (d. h. Wirksamkeit als prozentuale Veränderung). Der gesamte Ansatz ist damit räumlich explizit, berücksichtigt jedoch keine zeitliche Kompensation durch die
Fuchspopulation zwischen den einzelnen Modellschritten (d. h. zwischen M2 und M3). Die vorhergesagten Effekte entsprechen damit den erwarteten Effekten im Frühjahr unmittelbar nach Ende der Jagdzeit (bei einer konzentrierten Winterjagd im Studiengebiet).
Die Effektgröße bezieht sich auf Veränderungen im vorhergesagten Risiko einer Kunstgelegeprädation und kann daher keinesfalls absolut (d. h. im Sinne eines Effekts auf die Zielart), sondern nur relativ (d. h. im Vergleich der einzelnen Szenarien
untereinander) interpretiert werden.
3 Ergebnisse
Eine koordinierte Jagd auf der Gesamtfläche (alle angrenzenden Reviere; Szenario sim3, real3) hatte die höchste mittlere Effektgröße (reduziertes Prädationsrisiko) und die geringste Varianz auf der Fläche (Abb. 2). Eine Bejagung
im Randbereich angrenzend an die Zielfläche hatte die geringste mittlere Effektgröße und die größte Varianz auf der Fläche (Szenario sim2, real2). Die zentrale Jagd auf der Zielfläche erreichte mittlere Werte (Szenario sim1, real1).
Die Ergebnisse der beiden Ansätze „Simulation“ und „reales Gebiet“ glichen einander größtenteils, jedoch bei unterschiedlicher mittlerer Effektgröße. Im realen Untersuchungsgebiet lagen die mittleren Effektgrößen unter Berücksichtigung der landschaftlichen Variation sowie der
asymmetrischen Besitzverhältnisse deutlich unter denen der Simulation (Abb. 2b).
4 Diskussion
Die Prädatorenjagd und ihre Angemessenheit sind gesellschaftlich umstritten. Da dabei Tiere getötet werden, fordert das Tierschutzgesetz zudem einen „vernünftigen Grund“. Falls Prädatorenjagd zugunsten gefährdeter Tierarten angewendet wird, sollte dies darum auf eine Art und
Weise geschehen, die eine ausreichend hohe Wirksamkeit garantiert. Aber wie ist Wirksamkeit in diesem Zusammenhang zu beurteilen? Sie ist in der Praxis schwer zu bewerten, da sie selten quantifiziert wird (Doherty, Ritchie 2017; Lennox et al. 2018).
Der Gedanke, dass „ein toter Fuchs schon mal kein Beutetier mehr frisst“, dient deshalb so oder in ähnlicher Form häufig als Leitmotiv. Wie wir aber wissen, wird ein toter Fuchs schnell ersetzt (z. B. Newsome et al. 2014;
Lieury et al. 2015). Gemeint ist damit, dass eine Prädatorenkontrolle mit jagdlichen Mitteln nur dann für die Zielart effektiv wirksam sein kann, wenn es durch die Maßnahme zu einer geringeren Kontaktrate zwischen Beute und
Prädator kommt. Im einfachsten Fall wird dies durch eine jagdlich bedingte Senkung der Prädatorendichte erreicht. Sind die jagdlichen Eingriffe aber so strukturiert, dass die erlegten Tiere vor dem Zielzeitraum ausgeglichen werden (z. B. durch Einwanderung nicht-territorialer
Füchse) oder die Eingriffe räumlich an der falschen Stelle wirken, dann sind die Maßnahmen für den Artenschutz nicht wirksam.
In dieser Auswertung haben wir uns der Frage einer räumlichen Diskrepanz zwischen lokalen Effekten auf Ebene der Jagdreviere und der Zielflächen für den Artenschutz gewidmet. Aufgrund des deutschen Revierjagdsystems findet eine Bejagung oft dort statt, wo eine entsprechende
Motivation zur Fuchsbejagung oder wo entsprechende Ressourcen verfügbar sind, und damit nicht zwingend dort, wo es für die Zielart am relevantesten wäre (siehe z. B. Szenario real2, Abb. 2a).
Mithilfe der Modellkette konnten wir die räumlich stark begrenzte Beschaffenheit der herbeigeführten Effekte auf Fuchshäufigkeit und Prädationsrisiko visualisieren (Abb. 2a). Eine Bejagung in allen an die Zielfläche angrenzenden
Jagdrevieren (sim3, real3) erreichte konstant die größten Effekte auf der gesamten Zielfläche (vgl. Varianz um mittlere Effektgrößen in Abb. 2b), jedoch mit deutlich höherem jagdlichem Aufwand. Das bedeutet, dass diese Bejagungsform
auf der gesamten Zielfläche wirksam war. Zwar erreichte eine Bejagung in einzelnen Revieren am Rand der Zielfläche (sim2, real2) ebenfalls punktuell vergleichbar große Effekte (d. h. am Bejagungsschwerpunkt, Abb. 2a), jedoch
waren auf großen Teilen der Zielfläche nur sehr geringe oder keine Effekte zu erwarten (grüne Flächen in Abb. 2a).
In der Praxis bedeutet dies, dass eine Bejagung in einzelnen Randlagen – ungeachtet ihrer Intensität – auf einem erheblichen Teil der Zielfläche nicht wirksam ist. Eine höhere Varianz war auch bei einer zentralen Bejagung auf der Zielfläche (sim1, real1) zu
beobachten (Abb. 2b unten), jedoch stellt eine zentrale Bejagung in der Praxis möglicherweise eine kostengünstigere, wenngleich potenziell weniger wirksame Alternative dar.
Die erzielten Effektgrößen im realen Untersuchungsgebiet waren deutlich geringer als in der Simulation, auch wenn diese nicht vollständig miteinander vergleichbar sind. Dies ist auf die ungleichmäßige Verzahnung der Jagdreviergrenzen (z. B. aufgrund der Gegebenheiten des
Landbesitzes), auf Unterschiede in der landschaftlichen Ausstattung (und damit in der Fuchsdichte) und auf die ungleichmäßige Form der Zielfläche (die aber den Lebensraumansprüchen der Zielart entspricht) zurückzuführen. Wenngleich die Effektgrößen in unserem Ansatz nicht in den
tatsächlichen Reproduktionserfolg der Zielart übersetzt werden können, so zeigen die geringeren Effektgrößen im realen Gebiet doch die hohe Komplexität und die Grenzen der Prädatorenjagd auf großen Flächen im Kontext einer fragmentierten Waldlandschaft (z. B. im Vergleich zum
oft kleinräumigen Schutz bekannter Brutgebiete von Bodenbrütern im Offenland).
Eine gezielte Bejagung kann zwar eine lokale Reduktion der Fuchshäufigkeit erreichen (Kämmerle et al. 2019b; Porteus et al. 2019). Die notwendige jagdliche Intensität auf der
Fläche herbeizuführen, bedeutet in der Praxis jedoch einen hohen Aufwand, wie es z. B. beim großflächigen Raufußhuhn-Management in Großbritannien mit Berufsjägerinnen und -jägern der Fall ist. Konkret lassen sich die Herausforderungen an einem Beispiel verdeutlichen: Im realen
Untersuchungsgebiet im Schwarzwald liegt die erwartete Fuchsdichte (basierend auf Streifgebietsgrößen) bei bis zu 5 Tie-ren/km²; das bedeutet, dass diese bei einer jagdlichen Intensität von 5 Füchsen/km² abgeschöpft oder überschritten wird.
Die größten Effektgrößen werden mit einer koordinierten Jagd in allen Revieren erzielt, die an die Auerhuhn-Zielfläche angrenzen (sim3; real3); davon sind im Beispiel knapp 10 000 ha Revierfläche direkt betroffen. Demnach müssten auf dieser Fläche in der
Wintersaison zwischen Dezember und Anfang Februar mindestens ca. 500 Füchse erlegt werden. Das entspricht dem zwei- bis dreifachen der Jagdstrecke, die gegenwärtig mit einer stark heterogenen Verteilung der Bejagungsintensität erreicht wird.
Eine flächig intensive Bejagung über Jagdreviergrenzen hinweg ebenso wie eine zentral koordinierte Bejagung stellen damit vor allem eine enorme praktische Herausforderung dar, da diese von den Revierinhaberinnen und -inhabern selbst oder unter Einsatz von Berufsjägerinnen und
-jägern realisiert werden müsste, auch wenn die rechtlichen Voraussetzungen für eine zentrale Steuerung zu Naturschutzzwecken gegeben sind (z. B. § 36 Jagd- und Wildtiermanagementgesetz Baden-Württemberg).
Weiterhin ist neben der räumlichen Verteilung der Mortalität auch der Zeitpunkt ein wichtiger Faktor für die Wirksamkeit. Für Bodenbrüter soll meistens das Prädationsrisiko in der Reproduktions-zeit gesenkt werden. Unsere Untersuchungen zeigten allerdings, dass eine durch
gezielte Fuchsjagd herbeigeführte Reduktion innerhalb von ca. 10 Wochen nach Ende der Jagd schon nicht mehr nachzuweisen war und damit den für die Zielart relevantesten Zeitraum verfehlte (Kämmerle et al. 2019a, b). Eine Bejagung während der Reproduktionszeit der Zielart steht in unserem Beispiel in Baden-Württemberg aber in klarem Widerspruch zum Jagdrecht und zu dem darin enthaltenen Gedanken des Tierschutzes.
Prädatorenkontrolle kann zwar eine hochwirksame Maßnahme im Artenschutz sein, ist allerdings auch mit enormen Hürden (z. B. gesetzliche Restriktionen, Ressourcen, geringe Akzeptanz in der Bevölkerung, möglicherweise geringe Effektgrößen) verbunden. Es ist zu bedenken, dass
der Eingriff wahrscheinlich wirkungslos bleibt, wenn die Bejagung nicht räumlich und zeitlich auf die Ökologie der Zielart abgestimmt ist. Unsere Ergebnisse zeigen, dass eine Bejagung bestenfalls auf der gesamten für die Zielart relevanten Fläche koordiniert und mit
maximaler Intensität durchgeführt werden sollte. Eine koordinierte zentrale Bejagung auf der Zielfläche kann eine kostengünstigere Alternative mit potenziell geringerer Wirksamkeit darstellen. Im besten Fall wird die Wirksamkeit auch durch ein (Erfolgs)monitoring begleitet.
5 Literatur
↑
Baines D. (1996): The implications of grazing and predator management on the habitats and breeding success of black grouse Tetrao tetrix. Journal of Applied Ecology
33(1): 54 – 62.
↑
Baines D., Aebischer N.J., Macleod A. (2016): Increased mammalian predators and climate change predict declines in breeding success and density of Capercaillie
Tetrao urogallus, an old stand specialist, in fragmented Scottish forests. Biodiversity and Conservation 25(11): 2 171 – 2 186.
↑
Bino G., Dolev A. et al. (2010): Abrupt spatial and numerical responses of overabundant foxes to a reduction in anthropogenic
resources. Journal of Applied Ecology 47(6): 1 262 – 1 271.
↑
Braunisch V., Suchant R. (2006): Das Raufusshühner-Bestandesmonitoring der FVA. Berichte Freiburger Forstliche Forschung 64: 47 – 65.
↑
Chautan M., Pontier D., Artois M. (2000): Role of rabies in recent demographic changes in red fox (Vulpes
vulpes) populations in Europe. Mammalia 64(4): 391 – 410.
↑
Conner L.M., Morris G. (2015): Impacts of mesopredator control on conservation of mesopredators and their prey. PLOS ONE 10(9): e0137169.
↑
Coppes J., Ehrlacher J. et al. (2019): Dramatischer Rückgang der Auerhuhnpopulation Tetrao urogallus im Schwarzwald. Vogelwarte 57:
115 – 122.
↑
Côté I.M., Sutherland W.J. (1997): The effectiveness of removing predators to protect bird populations. Conservation Biology 11(2):
395 – 405.
↑
Doherty T.S., Ritchie E.G. (2017): Stop jumping the gun: A call for evidence-based invasive predator management. Conservation Letters 10(1):
15 – 22.
↑
Elmhagen B., Ludwig G. et al. (2010): Top predators, mesopredators and their prey: Interference ecosystems along bioclimatic
productivity gradients. Journal of Animal Ecology 79(4): 785 – 794.
↑
Elmhagen B., Rushton S.P. (2007): Trophic control of mesopredators in terrestrial ecosystems: Top-down or bottom-up? Ecology Letters 10(3):
197 – 206.
↑
Fahrig L. (2003): Effects of habitat fragmentation on biodiversity. Annual Review of Ecology, Evolution, and Systematics 34(1):
487 – 515.
↑
Fischer J., Lindenmayer D.B. (2007): Landscape modification and habitat fragmentation: A synthesis. Global Ecology and Biogeography 16(3):
265 – 280.
↑
Güthlin D., Storch I., Küchenhoff H. (2013): Landscape variables associated with relative abundance of generalist mesopredators. Landscape Ecology 28(9):
1 687 – 1 696.
↑
Güthlin D., Storch I., Küchenhoff H. (2014): Toward reliable estimates of abundance: Comparing index methods to assess the abundance of a mammalian predator. PLOS ONE
9(4): e94537.
↑
Huhta E., Helle P. et al. (2017): The effect of human-modified landscape structure on forest grouse broods in two landscape types. Ecosphere 8:
e01950.
↑
Jahren T., Storaas T. et al. (2016): Declining reproductive output in capercaillie and black grouse – 16 countries and
80 years. Animal Biology 66(3 – 4): 363 – 400.
↑
Kämmerle J.-L., Coppes J. et al. (2017): Range loss of a threatened grouse species is related to the relative abundance of a mesopredator.
Ecosphere 8(9): Article e01934.
↑
Kämmerle J.-L., Niekrenz S., Storch I. (2019a): No evidence for spatial variation in predation risk following restricted-area fox culling. BMC Ecology
19(1): 17.
↑
Kämmerle J.-L., Ritchie E.G., Storch I. (2019b): Restricted-area culls and red fox abundance: Are effects a matter of time and place? Conservation Science
and Practice 1(11): e115.
↑
Kämmerle J.-L., Storch I. (2019): Predation, predator control
and grouse populations: A review. Wildlife Biology 2019: wlb.00464.
↑
Kaphegyi T. (2002): Untersuchungen zum Sozialverhalten des Rotfuchses (Vulpes vulpes L.). Inaugural-Disseration. Albert-Ludwigs-Universität
Freiburg i. Br. Freiburg i. Br.: 109 S.
↑
Klaus S., Andreev V. et al. (1989): Die Auerhühner. 2. Aufl. Neue Brehm Bibliothek. Ziemsen Verlag. Wittenberg-Lutherstadt: 276 S.
↑
Kurki S., Linden H. (1995): Forest fragmentation due to agriculture affects the reproductive success of the ground-nesting black grouse Tetrao
tetrix. Ecography 18(2): 109 – 113.
↑
Kurki S., Nikula A. et al. (1998): Abundances of red fox and pine marten in relation to the composition of boreal forest
landscapes. Journal of Animal Ecology 67(6): 874 – 886.
↑
Kurki S., Nikula A. et al. (2000): Landscape fragmentation and forest composition effects on grouse
breeding success in boreal forests. Ecology 81(7): 1 985 – 1 997.
↑
Langgemach T., Bellebaum J. (2005): Prädation und der Schutz bodenbrütender Vogelarten in Deutschland. Vogelwelt 126(4): 259 – 298.
↑
Lennox R.J., Gallagher A.J. et al. (2018): Evaluating the efficacy of predator removal in a conflict-prone world. Biological Conservation
224: 277 – 289.
↑
Lieury N., Ruette S. et al. (2015): Compensatory immigration challenges predator control: An experimental evidence-based approach improves
management. The Journal of Wildlife Management 79(3): 425 – 434.
↑
Ludwig G.X., Alatalo R.V et al. (2010): Individual and environmental determinants of daily black grouse nest survival rates at variable predator densities. Annales
Zoologici Fennici 47: 387 – 397.
↑
Newsome T.M., Crowther M.S., Dickman C.R. (2014): Rapid recolonisation by the European red fox: How effective are uncoordinated and isolated control programs?
European Journal of Wildlife Research 60(5): 749 – 757.
↑
Nychka D., Furrer R. et al. (2017): Fields: Tools for spatial data. R package version 10.3. DOI: 10.5065/D6W957CT
↑
Pasanen-Mortensen M., Elmhagen B. (2015): Land cover effects on mesopredator abundance in the presence and absence of apex predators. Acta
Oecologica 67: 40 – 48.
↑
Pasanen-Mortensen M., Pyykönen M., Elmhagen B. (2013): Where lynx prevail, foxes will fail – Limitation of a mesopredator in Eurasia. Global Ecology and
Biogeography 22(7): 868 – 877.
↑
Pegel M. (2004): Fuchsbejagung – Sinn oder Unsinn. WFS-Mitteilungen 1/2004: 1 – 4.
↑
Perry D.A., Perry G.A. (2008): Improving interactions between animal rights groups and conservation biologists. Conservation Biology 22(1):
27 – 35.
↑
Porteus T.A., Reynolds J.C., McAllister M.K. (2019): Population dynamics of foxes during restricted-area culling in Britain: Advancing understanding through
state-space modelling of culling records. PLOS ONE 14(11): e0225201.
↑
Prugh L.R., Stoner C.J. et al. (2009): The rise of the mesopredator. Bioscience 59(9): 779 – 791.
↑
Reynolds J.C., Tapper S.C. (1996): Control of mammalian predators in game management and conservation. Mammal Review 26(2 – 3):
127 – 155.
↑
Ritchie E.G., Johnson C.N. (2009): Predator interactions, mesopredator release and biodiversity conservation. Ecology Letters 12(9):
982 – 998.
↑
Šálek M., Drahníková L., Tkadlec E. (2015): Changes in home range sizes and population densities of carnivore species along the natural to urban habitat gradient.
Mammal Review 45(1): 1 – 14.
↑
Schadt S., Revilla E. et al. (2002): Assessing the suitability of central European landscapes for the
reintroduction of Eurasian lynx. Journal of Applied Ecology 39(2): 189 – 203.
↑
Small M.F., Hunter M.L. (1988): Forest fragmentation and avian nest predation in forested landscapes. Oecologia 76(1): 62 – 64.
↑
Smith R.K., Pullin A.S. et al. (2010): Effectiveness of predator removal for enhancing bird populations. Conservation Biology 24(3):
820 – 829.
↑
Storch I. (1994): Habitat and survival of capercaillie Tetrao urogallus nests and broods in the Bavarian Alps. Biological Conservation 70(3):
237 – 243.
↑
Storch I. (1995): Annual home ranges and spacing patterns of capercaillie in central Europe. The Journal of Wildlife Management 59(2): 392 – 400.
↑
Storch I., Woitke E., Krieger S. (2005): Landscape-scale edge effect in predation risk in forest-farmland mosaics of central Europe. Landscape Ecology 20(8):
927 – 940.
↑
Suchant R., Braunisch V. (2008): Rahmenbedingungen und Handlungsfelder für den Aktionsplan Auerhuhn: Grundlagen für ein integratives
Konzept zum Erhalt einer überlebensfähigen Auerhuhnpopulation im Schwarzwald. Forstliche Versuchsanstalt Baden-Württemberg (FVA). Freiburg: 69 S.
↑
Wegge P., Rolstad J. (1986): Size and spacing of capercaillie leks in relation to social behavior and habitat. Behavioral Ecology and Sociobiology 19(6):
401 – 408.