Vera Luthardt, Oliver Brauner, Jenny Hammerich, Robert Probst, Corinna Schulz und Silke Finn Wachtel
Zusammenfassung
Der Klimawandel wirkt auf die wenigen noch wachsenden Moore ein, so dass die Frage besteht, inwieweit die Resilienz dieser
autochthonen Ökosysteme in all ihrer Vielfalt gestützt werden kann. Zur Beantwortung werden Dauerbeobachtungsreihen von weitgehend
ungestörten Mooren aus dem Biosphärenreservat Schorfheide-Chorin (Brandenburg) ausgewertet. Diese werden mit den Ergebnissen einer
Erfolgskontrolle wiedervernässter Waldmoore in Kontext gesetzt. Zur Einschätzung der Moorzustände wird ein neu entwickeltes
Indikatorensystem zur Bewertung moorspezifischer Biodiversität angewendet. Es wird zudem eine Abschätzung der Treibhausgasemissionen
nach der Treibhaus-Gas-Emissions-Standort-Typen(GEST)-Methodik vorgenommen und die potenzielle Torfneubildung betrachtet. Die Analysen
zeigen, dass das Puffervermögen wachsender Moore im Untersuchungsraum noch intakt ist und Störungen ohne Systemwechsel überwunden
werden. Die Vernässungsmaßnahmen waren durchweg erfolgreich und haben zu einer messbaren Revitalisierung geführt. Es wird auf die
dringende Notwendigkeit hingewiesen, heute alle noch weitestgehend naturnahen Moore in ihrem Wasserhaushalt bestmöglich zu
stabilisieren, um sie als wichtige Glieder der autochthonen Biodiversität mit allen ihren positiven Landschaftsfunktionen zu
erhalten.
Klimawandelanpassung – Klimawandelwirkung – Libellen – Monitoring – moorspezifische Biodiversität – moorspezifische Vegetation – MoorzustandAbstract
Climate change is affecting the few remaining mires that are still accumulating peat. The question thus arising is this: To what
extent can the resilience of these autochthonous ecosystems, in all their diversity, be enhanced? For this purpose, long-term
observation series of mostly undisturbed peatlands in the Schorfheide-Chorin Biosphere Reserve in the German regional state of
Brandenburg are evaluated. These are set in context with the findings of success monitoring of rewetted forest peatlands. A newly
developed indicator system for assessing mire-specific biodiversity is used to evaluate the state of the peatland. In addition,
greenhouse gas emissions are estimated using the GEST (greenhouse gas emissions habitat types) method and potential new peat formation
is considered. The analyses show that the buffering capacity of peat accumulating peatlands in the study area is still intact and that
disturbances can be overcome without changing the system. The waterlogging measures were consistently successful and led to a
measurable revitalisation. The article underscores the urgent need to stabilise the water balance of all peatlands that are still in a
near-natural state. This is vital in order to preserve them as important elements of autochthonous biodiversity with all their positive
landscape functions.
Climate change adaptation – Climate change impact – Dragonflies – Monitoring – Mire-specific biodiversity – Mire-specific vegetation – Peatland stateInhalt
1 Einführung
Naturnahe Moore und Ökosysteme auf Moorböden sind in den letzten Jahren verstärkt in den Fokus öffentlicher Aufmerksamkeit gerückt.
In erster Linie wird der Aspekt der klimaschädlichen Wirkung entwässerter Moore thematisiert. Entwässerte Moore emittieren große Mengen an
Treibhausgasen (THG) und tragen somit zum Klimawandel bei (Couwenberg et al. 2011; Tanneberger et al. 2021). So werden Moore – genauer gesagt Gebiete mit organischen Böden, von denen
sich in Deutschland 92 % im entwässerten Stadium befinden (Tiemeyer et al. 2020; Hofer, Köbbing 2021) – als„Klimakiller“ bezeichnet, denn enorme Mengen an Emissionen, im Jahr 2020
waren es 7,5 % der gesamten THG-Emissionen Deutschlands, resultieren aus der Umsetzung einst akkumulierten Kohlenstoffs aus diesen Böden
(UBA 2022). Naturnahe Moore emittieren THG als Methan, wirken jedoch in der Summe durch
beständige Kohlenstofffestlegung als Senken (Evans et al. 2021). Bei Wassermangel ändert sich
dies – ebenso wie in den genutzten Moorböden – und führt zu CO2-Emissionen in erheblichem Ausmaß (bis zu ca.
20 t CO2-Äquivalente ∙ ha−1 ∙ Jahr−1; abgeleitet aus
Treibhaus-Gas-Emissions-Standort-Typen[GEST]-Werten in Reichelt 2015).
Doch Moore sind weit mehr als ein Kohlenstoffspeicher. Obwohl ihr Flächenanteil in früheren Zeiten in Deutschland nur ca. 5 % betrug
(Bonn et al. 2015), hat sich nach der Eiszeit eine sehr moorspezifische Lebewelt etabliert,
die einen erheblichen Anteil der autochthonen Biodiversität über alle ihre Ebenen hinweg ausmacht. Succow (2001) unterscheidet nach Hydrologie, Trophiestatus und Säure-Basen-Verhältnissen 23 landschaftsökologische
Moortypen. Diese differenzieren sich weiter durch die Höhenlage, die Verzahnung verschiedener Moortypen, das mosaikartige Nebeneinander
von Sukzessionsstadien und Vegetationsstrukturen, die individuelle Genese und die Kontinuität über die Zeit. Alle diese Komponenten
bedingen eine Vielzahl an Biozönosen und verleiten zu dem Satz: Kein Moor ist wie das andere.
In diesem Kontext ist auch die Bedeutung der Feuchtgebiete und naturnahen Moore im Zuge der Klimawandelanpassung hervorzuheben. Sie
sind Stabilisatoren des Landschaftswasserhaushalts, Hochwasserpuffergebiete, Feuchtluftsenken, Kühlungsräume und Rückzugsräume für diverse
Tierarten in heißen Perioden (Luthardt 2014). Diese Funktionen in der Landschaft spielen eine
zunehmende Rolle für Strategien der Anpassung von Regionen an veränderte Wetterbedingungen einschließlich des Abpufferns von
Extremereignissen.
Bei in Mooren natürlich vorkommenden Pflanzenarten wird zwischen moorspezifischen, d. h. nur in naturnahen Mooren vorkommenden Arten
und moortypischen Arten, d. h. Arten mit einer engen Bindung an Feuchtgebiete unterschieden (zugeordnete Pflanzenartenlisten siehe bspw.
für das Land Brandenburg Luthardt, Klawitter 2014).
Da heute nur noch maximal 5 % aller ehemaligen Moorflächen in einem naturnahen, d. h. lebensraumspezifischen Zustand vorhanden sind
(Bonn et al. 2015), sind Moore nach § 30 Bundesnaturschutzgesetz (BNatSchG) und auch in den
Landesnaturschutzgesetzen per se gesetzlich geschützte Biotope. Ein Großteil der moorspezifischen Arten steht auf den Roten Listen, z. B.
im Land Brandenburg 72 % der Gefäßpflanzenarten, 100 % der Libellen- und 59 % der Spinnenarten (Luthardt, Zeitz 2014). Obwohl oft von einer relativen Artenarmut der Moore gesprochen wird, bezieht sich dies vor allem auf
Blütenpflanzen. Beispielsweise gibt es bei den Insekten eine sehr reiche, oft winzige und unscheinbare Fauna, die in großen Teilen noch
nicht genauer untersucht ist (Barndt 2012SCB
2021).
Sieben der nach Fauna-Flora-Habitat(FFH)-Richtlinie geschützten Lebensraumtypen gehören zu Mooren und weitere 15 sind oft mit ihnen
vergesellschaftet. Somit bedeutet der vordringliche Schutz der einheimischen Biodiversität, einen starken Schwerpunkt auf die noch
naturnahen Moore zu lenken, wie es der International Congress for Conservation Biology 2021 () wieder eindringlich gefordert hat.
Hauptgefährdungsfaktor für Moorökosysteme ist eine kontinuierliche Wassermangelsituation (Landgraf 2007; Trepel et al. 2017). Eine anhaltende
Entwässerung nimmt auch den ungenutzten Mooren durch die Veränderung der Bodeneigenschaften (Degradierung des Akrotelms als der
wasserdurchleitenden, aktiven obersten Torfschicht; Sackung und Verdichtung des Katotelms als des darunter liegenden Hauptwasserspeichers;
Veränderung des Mikroklimas durch ausbleibende Verdunstung; Auteutrophierungseffekte durch Mineralisierung des oberflächennahen Torfs) die
Fähigkeit der Selbstregulation. Damit sinkt die Resilienz gegenüber klimatischen Veränderungen (Bonn et
al. 2016; Luthardt, Wichmann 2016). Deshalb verweisen viele Publikationen darauf,
dass Moore generell auf Grund der zu erwartenden Veränderungen des Landschaftswasserhaushalts zu den stark durch den Klimawandel
gefährdeten Ökosystemen gehören (Petermann et al. 2007; Beierkuhnlein et al. 2014; Hauck et al. 2019). Dies ist jedoch zu
differenzieren. Je nach Klimaregion, Veränderungen des Gebietswasserhaushalts, hydrologischem Moortyp und Einbettung ins Gelände werden
vermutlich sehr verschiedenartige Entwicklungen eintreten (Luthardt, Zeitz 2014; Hauck et al. 2019).
Der Blick auf die Moorarten zeigt, dass eine Vielzahl zu hauptsächlich boreal verbreiteten Arten gehört, die u. U. direkt auf den
Anstieg der Temperatur reagieren könnten. Nach Hauck et al. (2019) spielt die
Temperaturerhöhung jedoch bisher noch keine Rolle in intakten Mooren. Moore stellen Mikroklimaräume dar, die ein gewisses Puffervermögen
gegenüber der Umgebungstemperatur aufweisen. Unbekannt ist, wo der tipping point liegt. Nach Pompe
et al. (2011) würde bei der Projektion von Klimawandelszenarien auf die Verbreitungsgebiete von Arten nach ihren bisherigen
klimatischen Hüllen bspw. die Sumpfdotterblume (Caltha palustris) bei einer angenommenen durchschnittlichen Temperaturerhöhung von
2 K nur sehr kleinflächig in Deutschland verschwinden – bei einer Erhöhung um 4 K wäre der Verlust jedoch wesentlich
dramatischer.
Neben diesen direkten klimatischen Faktoren spielen auch Sekundäreffekte, wie eine zunehmende Eutrophierung durch beschleunigte
Prozesse der Umsetzung anfallender rezenter toter Biomasse und die Düngung über Lufteinträge, eine nicht zu unterschätzende Rolle für die
nährstoffärmeren Moorökosysteme, was einen zunehmenden Verlust nicht konkurrenzfähiger Pflanzenarten und damit einen Wandel des Struktur-
und Nahrungsangebots für alle vergesellschafteten Pilz- und Tierarten befürchten lässt (Hanspach et al.
2013; Hauck et al. 2019).
Um nachzuvollziehen, wie sich die beschriebenen Effekte auf naturnahe Moore auswirken und mit welchen Maßnahmen den möglichen
Entwicklungen entgegengewirkt werden kann, sind Dauerbeobachtungsreihen erforderlich. Es werden Fallbeispiele für das Biosphärenreservat
(BR) Schorfheide-Chorin vorgestellt, die Teil der Ökosystemaren Umweltbeobachtung (ÖUB) der drei Biosphärenreservate Brandenburgs (http://maps18.hnee.de/oeub/index.html) sind. Im BR Schorfheide-Chorin
wurden zudem bereits vielfältige Renaturierungsprojekte in Mooren durchgeführt. Einige davon sind Teil des Brandenburger
Waldmoorschutzprogramms. Ziel dieses Programms ist der Schutz der naturnahen und die Wiederherstellung degradierter Moore (MLUL 2016). Von 2017 bis 2022 erfolgte eine Erfolgskontrolle der Renaturierungsmaßnahmen (Hammerich et al. 2022b), aus der hier Teilergebnisse vergleichend hinzugezogen werden.
2 Methodisches Vorgehen und untersuchte Moorstandorte
Das BR Schorfheide-Chorin ist gekennzeichnet durch eine vielfältige Ausstattung mit diversen Moortypen in einem erheblichen
Flächenanteil (Abb. 1). Die ÖUB beobachtet sechs naturnahe Moore. Diese sind überwiegend als
oligotroph- bzw. mesotroph-saure Kessel- oder Verlandungsmoore zu charakterisieren (Tab. 1).
Detailinformationen zum Untersuchungsprogramm für Moorökosysteme im Rahmen der ÖUB sind dem Methodenkatalog (Luthardt et al. 2017) zu entnehmen. Seit 1999 werden neben Erfassungen abiotischer Faktoren wie
Pegelstand, Gesamtkohlenstoff (Ct), Gesamtstickstoff (Nt) und pH im Oberboden auch detaillierte Vegetationsanalysen im drei- bis
sechsjährigen Untersuchungsintervall durchgeführt: Gesamtartenliste, Vegetationstransekte und Vegetationsaufnahmen nach Braun-Blanquet
(Tab. A im Online-Zusatzmaterial). Weiterhin wurden 2002
und 2018 Vegetationszonierungen kartiert. Dafür wurden die Moorflächen in homogene Vegetationseinheiten nach floristischer Dominanz,
ökologischen Standortbedingungen und Physiognomie aufgeteilt. Pro Vegetationseinheit wurden alle Pflanzenarten mit Deckungen erfasst.
Durch Anwendung des Vegetationsformenkonzepts (Clausnitzer, Succow 2001; Koska et al. 2001; Roth, Succow 2001) konnten
flächenscharfe Wasserstands-, Trophie- und Säure-Basen-Verhältnisse je Moor abgeleitet werden. Faunistische Untersuchungen erfolgen im
Rahmen der ÖUB alle drei Jahre zum Libellenvorkommen. Um die Beobachtungen in einen klimatischen Rahmen zu setzen, werden die
Wetterdaten der nächstgelegenen Klimastation Angermünde (DWD 2022) als Referenzstation
verwendet.
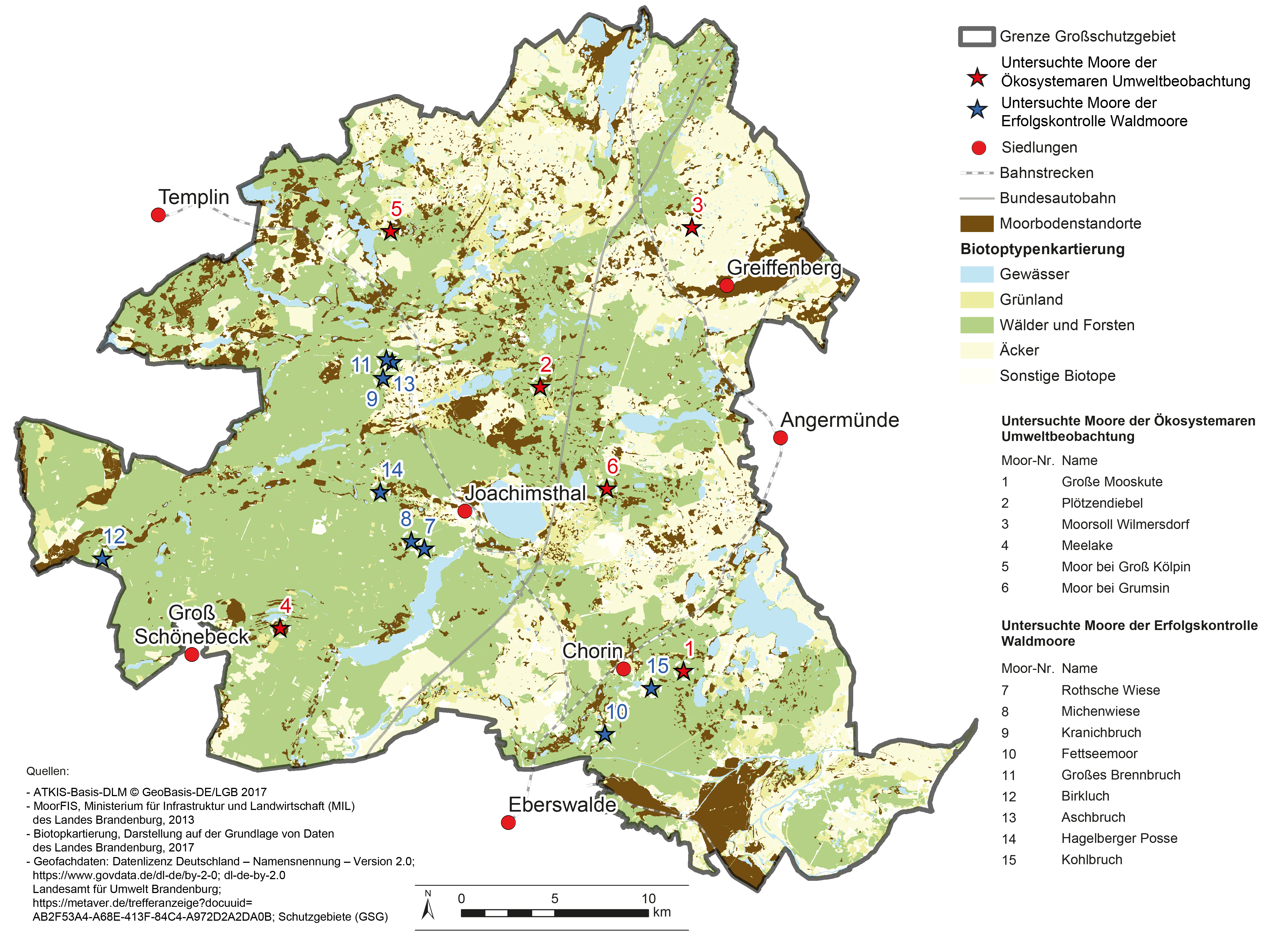
Abb. 1: Moorböden im Biosphärenreservat Schorfheide-Chorin mit Verortung der untersuchten Moore.
Fig. 1: Peat soils in the Schorfheide-Chorin Biosphere Reserve with the locations of the peatlands studied.
|
Moor-Nr.
|
Name
|
Größe (ha)
|
Hydrogenetischer Moortyp
|
Säure-Basen-Stufe
|
Trophie
| |
ÖUB-Moore |
1
| Große Mooskute | 1,0 | Kesselmoor | Sauer | Oligotroph | 1999 |
2
| Plötzendiebel | 17,0 | Kessel-Verlandungsmoor | Sauer | Oligotroph | 1999 |
3
| Moorsoll Wilmerdorf | 0,6 | Kesselmoor | Sauer | Mesotroph | 1999 |
4
| Meelake | 44,2 | Verlandungsmoor | Sauer | Oligotroph/mesotroph | 2001 |
5
| Moor bei Groß Kölpin | 8,8 | Versumpfungs- über Verlandungsmoor | Sauer | Mesotroph | 2018 |
6
| Moor bei Grumsin | 2,0 | Kesselmoor | Sauer | Oligotroph/mesotroph | 2018 |
Renaturierte Waldmoore |
7
| Rothsche Wiese | 5,4 | Verlandungsmoor | Subneutral | Eutroph | 2010 |
8
| Michenwiese | 2,0 | Verlandungsmoor | Subneutral | Eutroph | 2010 |
9
| Kranichbruch | 3,0 | Versumpfungsmoor | Subneutral | Eutroph | 2006 |
10
| Fettseemoor | 9,6 | Verlandungsmoor | Sauer/subneutral | Eutroph/mesotroph | 1991 |
11
| Großes Brennbruch | 4,9 | Verlandungsmoor | Sauer | Mesotroph | 2006 |
12
| Birkluch | 6,1 | Durchströmungsmoor | Subneutral | Eutroph | 2000 |
13
| Aschbruch | 1,1 | Versumpfungsmoor | Subneutral | Eutroph | 2006 |
14
| Hagelberger Posse | 23,0 | Versumpfungsmoor | Sauer | Mesotroph | 2008 |
15
| Kohlbruch | 4,0 | Versumpfungsmoor | Subneutral | Eutroph | 1995 |
* Bei renaturierten Mooren Jahr der Maßnahmenumsetzung; Namen zu den Moor-Nr. siehe Abb. 1 |
Tab. 1: Kurzbeschreibung der untersuchten Moore: Moore in der Ökosystemaren Umweltbeobachtung (ÖUB) und renaturierte und
evaluierte Waldmoore im Biosphärenreservat Schorfheide-Chorin.
Table 1: Brief description of the peatlands studied: Peatlands covered by the ecosystemic environmental monitoring scheme
(“Ökosystemare Umweltbeobachtung – ÖUB”), and restored and evaluated forest peatlands in the Schorfheide-Chorin Biosphere
Reserve.
Die Kartierungen zur Erfolgskontrolle der Waldmoorrenaturierungen erfolgten von 2018 bis 2020. Neun der untersuchten Moore liegen im
BR Schorfheide-Chorin. An allen Standorten wurden wasserbauliche Maßnahmen durchgeführt, d. h., der Abfluss aus dem Moor wurde mittels
Lehmplomben außerhalb des Moors verschlossen und/oder die Gräben innerhalb des Moores wurden verfüllt. Bei der Kartierung wurden die
Moorflächen in homogene Vegetationseinheiten aufgeteilt. Pro Vegetationseinheit wurden alle Pflanzenarten mit Deckung sowie die obersten
30 cm des Bodens (Torfart, Zersetzungsgrad, Farbe, Beimengungen) erfasst, um die Torfdegradierung und etwaige neue Torfbildung zu
dokumentieren. Auf Grundlage der Vegetationseinheiten wurde das Vegetationsformenkonzept (Succow,
Joosten 2001) angewendet und die zuvor genannten moorrelevanten Standortindikatoren wurden abgeleitet. Der jeweilige
Moor-zustand vor Renaturierung wurde durch Analyse von Altdaten analog ermittelt.
Für den Vergleich der Feuchteverhältnisse auf der Gesamtmoorfläche über die Zeitreihen wurde der Flächenanteil der unterschiedlichen
Wasserstufen als Ausdruck der Jahresmediane des Moorwasserstands (Succow, Joosten 2001; Tab. B
im Online-Zusatzmaterial) gemittelt. Der Wert fünf
entspricht einem flurgleichen Wasserstand auf der gesamten Moorfläche.
Als weitere Vergleichsbasis in der Auswertung wurde für alle Moore die „moorspezifische Biodiversität“ zu den verschiedenen
Zeitpunkten mit dem Indikatorensystem nach Hammerich et al. (2022a) bestimmt. Dazu wurden als
Teilindikatoren die moorspezifische Flora, die Habitatvielfalt und -vernetzung und der Ökosystemzustand als Bodendegradierungsgrad und
Wasserstufe über ein fünfstufiges Punktesystem bewertet. Die Einzelbewertungen wurden summiert, so dass ein Maximalwert von 15 für eine
„sehr gute moorspezifische Biodiversität“ vergeben werden konnte (für weitere Abstufungen und Informationen zum Indikatorensystem siehe
Abschnitt 3 und Abb. A, B im Online-Zusatzmaterial).
Die THG-Emissionen aus den Mooren wurden basierend auf den Vegetationsformenanalysen mithilfe der GEST nach Couwenberg et al. (2011), Spangenberg (2011) und
Reichelt (2015) berechnet. Die Fläche, auf der aktives Torfwachstum stattfindet, wurde für
die jeweiligen Vegetationseinheiten anhand der Oberbodenansprache abgeschätzt. Torfbildung wurde angenommen, wenn flurnahe Wasserstände
vorherrschten, torfbildende Vegetation wuchs und diese den obersten abgrenzbaren Horizont durchwurzelte (bei Seggen) oder eine neue
abgrenzbare Ablage abgestorbener Moose vorgefunden wurde. Alle hier betrachteten Moore sind in Tab. 1 aufgeführt, Steckbriefe zu den einzelnen Mooren mit genauer Verortung und Beschreibung sind in Luthardt et al. (2022) zu finden.
3 Entwicklung der naturnahen Moore der ökosystemaren Umweltbeobachtung im Biosphärenreservat Schorfheide-Chorin
Im langjährigen Mittel von 1961 bis 1990 betrug die Jahresmitteltemperatur in der Region 8,3 °C und die Jahressumme des Niederschlags
532 mm. Im Zeitraum 1998 – 2021 lagen die Jahresmitteltemperaturen (bis auf das Jahr 2007) über diesem langjährigen Mittel mit
Abweichungen von bis zu 2,6 K. Gegenüber dem lang-jährigen Mittel der jährlichen Niederschlagssumme hat sich die mittlere jährliche
Niederschlagssumme im ÖUB-Zeitraum kaum verändert. Es lassen sich zwei größere Phasen ausmachen: 2007 – 2012 mit feuchten Jahren; seit
2013 trockene Phase mit Jahressummen deutlich unterhalb des langjährigen Mittels (nur 2017 sehr feucht; Abb. 2). Die Phänologiedaten der Meldestationen im BR Schorfheide-Chorin (Abb. C im Online-Zusatzmaterial) zeigen eine Verlängerung der
Vegetationsperiode um 24 Tage im ÖUB-Zeitraum (1998 – 2021) gegenüber dem langjährigen Referenzzeitraum (1961 – 1990). Vor allem im Herbst
verlängerten sich die Phänophasen.
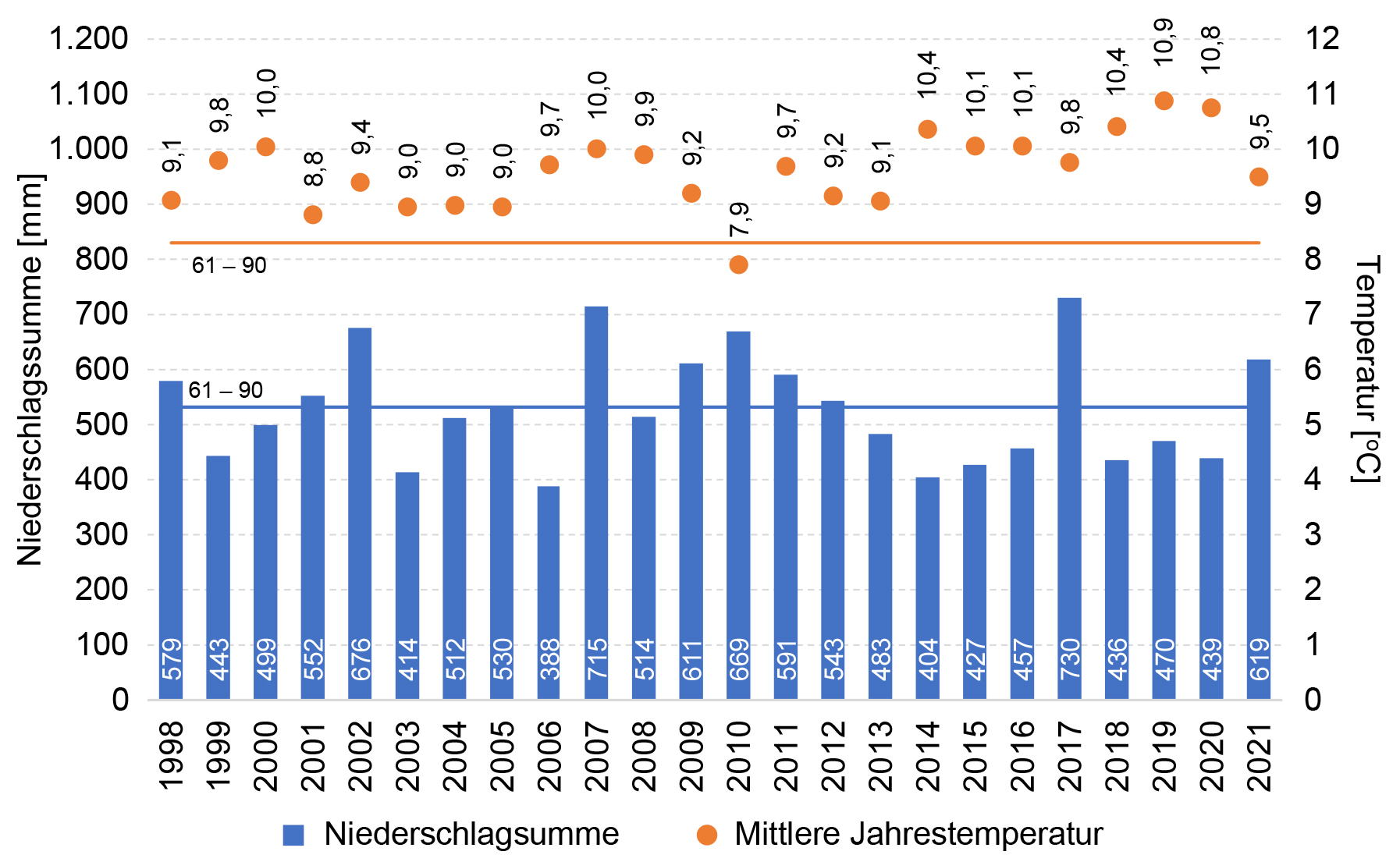
Abb. 2: Jahresmitteltemperaturen und Jahresniederschlagssummen der Station Angermünde für den Zeitraum 1998 – 2021 und das
langjährige Mittel (1961 – 1990) der Station.
Fig. 2: Annual mean temperatures and sum of annual precipitation at Angermünde station for the 1998 – 2021 period and the
long-term mean (1961 – 1990) of the station.
Alle in der ÖUB untersuchten Moore wiesen bei der Erstaufnahme 1999/2000 oberflächennah nicht degradierte schwach zersetzte Torfe auf
(Luthardt et al. 2005), die auf weitgehend kontinuierliche, wassergesättigte Bedingungen in
der nahen Vergangenheit hinweisen.
Alle erfassten moorspezifischen Gefäßpflanzenarten sind in Tab. C im Online-Zusatzmaterial aufgelistet. Die interessanten zyklischen Fluktuationen der Pflanzen- und Libellenarten (siehe
ausgewählte Arten und Lebensräume in Abb. 3, Abb. 4, 5) in Abhängigkeit von der Witterung und den damit einhergehenden
Veränderungen des Wasserhaushalts sind in den Steckbriefen für die einzelnen Moore von Luthardt et al.
(2022) dargestellt.

Abb. 3: Die in Brandenburg stark gefährdete und in den letzten Jahren zunehmend seltenere Gefleckte Heidelibelle (Sympetrum
flaveolum; hier ein Weibchen bei großer Hitze in der so genannten Obeliskhaltung) wurde in den Jahren 2019 und 2021
vereinzelt bodenständig im Moor bei Groß Kölpin gefunden.
Fig. 3: The yellow-winged darter (Sympetrum flaveolum), which is highly endangered in Brandenburg and has become
increasingly rare in recent years, was found sporadically domestic at the “Moor bei Groß Kölpin” peatland in 2019 and 2021. The
female here is in the so-called “obelisk posture” during high heat.

Abb. 4: Männchen der Östlichen Moosjungfer (Leucorrhinia albifrons), die im Moorsee des Plötzendiebels über viele Jahre
eines der größten Vorkommen Nordostdeutschlands besaß.
Fig. 4: Male of the dark whiteface (Leucorrhinia albifrons), which had one of the largest populations in northeast
Germany in the central lake of the “Plötzendiebel” peatland for many years.

Abb. 5: Lagg des Moors bei Grumsin, Lebensraum u. a. der Großen Moosjungfer (Leucorrhinia pectoralis). Im Vordergrund ist
ein größerer Bestand der Sumpf-Calla (Calla palustris) zu sehen.
Fig. 5: Lagg of the “Moor bei Grumsin” as a habitat of, amongst others, the large white-faced darter (Leucorrhinia
pectoralis). In the foreground, a larger population of the marsh calla (Calla palustris).
Ein Vergleich des Biodiversitätswerts je Moor und Zeitreihe ergab eine überwiegend gute moorspezifische Biodiversität, im Fall von
Plötzendiebel und Großer Mooskute sogar eine sehr gute moorspezifische Biodiversität (Tab. 2, Tab. D im Online-Zusatzmaterial). Während die Indexwerte für die Große Mooskute und den Plötzendiebel konstant blieben bzw. sich sogar
verbesserten, verringerten sich die Werte von Moorsoll und Meelake zwischen den beiden Erhebungen leicht. Die Abnahmen sind
vor allem auf die Bewertung der Artendiversität, d. h. Schwankungen in der Anzahl moorspezifischer Pflanzenarten zurückzuführen (Abb. D im
Online-Zusatzmaterial). Für alle Moore ist hinsichtlich
ihrer Entwicklung eine natürliche Dynamik zu erkennen, die stark an den Wasserstand gekoppelt ist. Dieser wird weitgehend durch das
Witterungsgeschehen beeinflusst. Jedoch zeigen sowohl Große Mooskute als auch Plötzendiebel bisher ein hohes Puffervermögen gegenüber den
vergangenen, z. T. extremen Trockenphasen. Bei Moorsoll und Meelake waren die Schwankungen im Beobachtungszeitraum größer. Die beiden
durch natürliche Einflussfaktoren (den Biber – Castor fiber) wiedervernässten Moore bei Groß Kölpin und Grumsin entwickeln
zunehmend wieder Moorqualitäten.
4 Entwicklung renaturierter Moore im Biosphärenreservat Schorfheide-Chorin
Im Ergebnis der umgesetzten Renaturierungsmaßnahmen zeigt die Erfolgskontrolle eine positive Entwicklung der Wasserstände, der
moorspezifischen Biodiversität, der Torfbildung und auch der Reduktion an freigesetzten THG-Emissionen in allen untersuchten Mooren. Die
Werte des Vorher-nachher-Vergleichs können Tab. 2 entnommen werden.
|
|
Moor-Nr.
|
Mittlere Wasserstufe
|
Moorspezifische Biodiversität
|
Torfbildung (% der Gesamtmoorfläche)
|
THG (t CO2-Äq. ∙ ha−1 ∙ a−1)
|
|
EA
|
WA/EK
|
EA
|
WA/EK
|
EA
|
WA/EK
|
EA
|
WA/EK
|
ÖUB-Moore | 1 | 4,9 | 5,0 | 14 | 14 | 80 | 100 | 7,2 | 6,4 |
2 | 4,3 | 5,1 | 14 | 15 | 78 | 93 | 7,3 | 7,2 |
3 | 3,1 | 3,9 | 12 | 11 | 50 | 50 | 14,7 | 17,3 |
4 | 4,1 | 3,8 | 14 | 12 | 59 | 39 | 10,9 | 8,1 |
| 4,7 | K. A. | 12 | K. A. | 55 | K. A. | 5,7 | K. A. |
| 5,0 | K. A. | 12 | K. A. | 50 | K. A. | 5,6 | K. A. |
REW | 7 | 2,3 | 4,5 | 6 | 9 | 0 | 72 | 25,5 | 9,3 |
8 | 2,8 | 3,0 | 4 | 5 | 0 | 0 | 22,0 | 16,4 |
9 | 5,0 | 5,2 | 10 | 12 | 50 | 57 | 4,2 | 3,3 |
10 | 4,7 | 5,1 | 15 | 15 | 77 | 88 | 7,9 | 4,5 |
11 | 3,1 | 5,0 | 9 | 13 | 4 | 99 | 15,3 | 3,3 |
12 | 3,0 | 3,8 | 4 | 6 | K. A. | K. A. | 19,5 | 7,7 |
13 | 3,0 | 4,4 | 5 | 9 | 0 | 19 | 9,3 | 7,9 |
14 | 4,9 | 5,0 | 13 | 14 | 94 | 100 | 3,5 | 3,7 |
15 | 3,0 | 4,9 | 5 | 10 | 0 | 94 | 12,5 | 7,0 |
* Moor-Nr. 5 und 6: Erstaufnahme nach ÖUB-Methode (Moor) im Jahr 2018; Namen zu den Moor-Nr.
siehe Abb. 1EA = Erstaufnahme (ÖUB-Moore: 1999 – 2002, REW-Moore: 1991 – 2010); EK = Erfolgskontrolluntersuchung für REW-Moore
(2018 – 2020); WA = Wiederholungsaufnahme für ÖUB-Moore (2018); K. A. = keine Angabe; ÖUB = Ökosystemare Umweltbeobachtung;
REW = renaturierte und evaluierte Waldmoore; THG = Treibhausgase |
Table 2: Area-weighted mean water table, index of mire-specific biodiversity according to Hammerich et al. (2022a), estimated area of peat formation and average greenhouse gas emissions (GHG) per ha and year
estimated according to GEST approach (Couwenberg et al. 2011; Spangenberg 2011; Reichelt 2015) for peatlands
covered by the ecosystemic environmental monitoring scheme (“Ökosystemare Umweltbeobachtung – ÖUB”), and for forest peatlands
covered by success monitoring (REW).
Alle untersuchten Moore weisen deutlich gestiegene Wasserstände auf (siehe beispielhaft Abb. 6). Im Mittel aller Moore stiegen die Wasserstände bzw. Wasserstufen von 3,5 auf 4,5. Auf einigen Mooren entstand auch
partieller Überstau, fünf Moore zeigen im Mittel flurgleiche Wasserstände an (Mittelwert 5). Die moorspezifische Biodiversität mit einer
bestmöglichen Ausprägung von 15 Punkten ist im Mittel aller Moore von 7,9 auf 10,3 Punkte angestiegen. Auch hier reagierten alle neun
Moore positiv. Vier Moore haben schon eine gute bis sehr gute Bewertung erhalten, nur zwei Moore weisen trotz Wasseranstiegs noch eine
geringe moorspezifische Biodiversität auf. Potenzielle Torfbildung fand vor Renaturierung im Mittel auf 28 % und nach Renaturierung auf
66 % der Moorfläche statt. Nur bei einem Moor war noch keine initiale Torfbildung zu beobachten. Durch die Renaturierungsmaßnahmen sanken
bei Schätzung mittels GEST auf allen neun Mooren die jährlichen THG-Emissionen pro ha erheblich. Insgesamt werden in der Summe jährlich
ca. 285 t CO2-Äquivalente auf den neun untersuchten Mooren eingespart.

Abb. 6: Das Zentrum des renaturierten Waldmoors Großes Brennbruch: a) vor (im Jahr 2006) und b) nach der Renaturierung im
Jahr 2018. Durch die wasserbaulichen Maßnahmen konnte der Wasserstand angehoben werden. Die jungen Sandbirken (Betula
pendula) sind abgestorben. Es hat sich ein nasser Bestand aus Sumpf-Blutaugen (Comarum palustre) und Flatterbinsen
(Juncus effusus) mit Torfmoosen (u. a. Sphagnum fallax) entwickelt.
Fig. 6: The centre of the restored forest peatland “Großes Brennbruch”: a) before (in 2006) and b) after restoration in 2018.
The water level was raised by hydraulic measures and the young silver birch trees (Betula pendula) are dead. A wet stand
comprising Comarum palustre, Juncus effusus and peat mosses (e. g. Sphagnum fallax) has developed.
5 Resümee und Schlussfolgerungen
Die Ergebnisse zeigen, dass naturnahe Moore ein hohes Puffervermögen gegenüber den bisherigen klimatischen Veränderungen aufweisen,
wenn sie über einen weitgehend ungestörten Wasserhaushalt verfügen. Austrocknungsphasen können überstanden werden, wenn danach wieder
Auffüllungsphasen folgen. Wo die tipping points des mikroklimatischen Puffervermögens bei zunehmendem Anstieg der Durchschnitts- und
Wintertemperaturen für einzelne Arten liegen, ist derzeit nicht abzuschätzen.
Der einzige Weg, den negativen Auswirkungen des Klimawandels entgegenzuwirken, ist die Renaturierung der degradierten Moorstandorte
mit dem Ziel, einen möglichst naturnahen Zustand und damit die Fähigkeit zur Selbstregulation wiederherzustellen. Dabei sind insbesondere
drei Faktoren entscheidend:
● Wiederherstellung flurnaher Wasserstände und des ursprünglichen Wasserregimes,
● Reetablierung einer moortypischen bzw. -spezifischen, torfbildenden Vegetation,
● Regeneration des Oberbodens mit Einsetzen neuen Torfwachstums.
Die Ergebnisse des Vorher-nachher-Vergleichs von Renaturierungen zeigen deutlich, dass es möglich ist, essenzielle Faktoren, die zur
Resilienz der Moore im Klimawandel beitragen, zu begünstigen und in Richtung naturnaher Verhältnisse zu entwickeln (siehe auch Luthardt et al. 2021 zu renaturierten Mooren im Stechlinsee-Gebiet). Es muss heute der
Handlungsschwerpunkt gesetzt werden, alle ungenutzten, in irgendeiner Form anthropogen entwässerten Moore zu vernässen. Nur so können auch
degradierte Moore wieder hinreichend resilient für die Herausforderungen der klimatischen Veränderungen gemacht und Lebensraumqualitäten
für autochthone Arten geschaffen werden. Die Vermeidung von THG-Emissionen ist ein zusätzlicher Anreiz für die schnelle Umsetzung von
Renaturierungsmaßnahmen. Spezifizierte Handlungsanleitungen sind u. a. über https://dss-wamos.de für Waldniedermoore abrufbar oder in Leitfäden für die einzelnen Bundesländer zu finden (z. B. LUA 2004; BLfU 2010).
6 Literatur
↑
Barndt D. (2012): Beitrag zur Kenntnis der Arthropodenfauna der Zwischenmoore Butzener Bagen, Trockenes Luch und Möllnsee im NSG
Lieberoser Endmoräne (Land Brandenburg). Märkische Entomologische Nachrichten 14: 147 – 200.
↑
Beierkuhnlein C., Jentsch A. et al. (Hrsg.) (2014): Auswirkungen des Klimawandels auf Fauna, Flora und
Lebensräume sowie Anpassungsstrategien des Naturschutzes. Naturschutz und Biologische Vielfalt 137: 484 S.
↑
BLfU/Bayerisches Landesamt für Umweltschutz (Hrsg.) (2010): Moorrenaturierung kompakt – Handlungsschlüssel für die Praxis. BLfU. Augsburg:
41 S.
↑
Bonn A., Allott T. et al. (2016): Peatland restoration and ecosystem services: An introduction. In: Bonn A., Allott T. et al.
(Hrsg.): Peatland restoration and ecosystem services: Science, policy and practice. Cambridge University Press. Cambridge:
1 – 16.
↑
Bonn A., Berghöfer A. et al. (2015): Klimaschutz durch Wiedervernässung von kohlenstoffreichen Böden.
In: Naturkapital Deutschland – TEEB DE (Hrsg.): Naturkapital und Klimapolitik. Technische Universität Berlin, Helmholtz-Zentrum für
Umweltforschung – UFZ. Berlin, Leipzig: 125 – 145.
↑
Braun-Blanquet J. (1964):
Pflanzensoziologie. Grundzüge der Vegetationskunde. Springer. Berlin: 631 S. DOI: 10.1007/978-3-7091-8110-2
↑
Clausnitzer U., Succow M. (2001): Vegetationsformen der Gebüsche und Wälder. In: Succow, M., Joosten H.
(Hrsg.): Landschaftsökologische Moorkunde. 2. Aufl. E. Schweizerbart'sche Verlagsbuchhandlung. Stuttgart: 161 – 170.
↑
Couwenberg J., Thiele A. et al. (2011): Assessing greenhouse gas emissions from peatlands using vegetation as a proxy. Hydrobiologia 674: 67 – 89. DOI:
10.1007/s10750-011-0729-x
↑
DWD Datenservice (2022): CDC-Portal des Deutschen Wetterdienstes. https://cdc.dwd.de/portal/ (aufgerufen am 2.5.2022).
↑
Evans C., Peacock M. et al. (2021): Overriding water table control on managed peatland greenhouse gas emissions. Nature 593 (7.860): 1 – 7. DOI:
10.1038/s41586-021-03523-1
↑
Hammerich J., Dammann C. et al. (2022a): Assessing mire-specific biodiversity with an indicator based
approach. Mires and Peat 28(32): 1 – 29.
↑
Hammerich J., Luthardt V., Zeitz J. (2022b): Moorschutz in den Wäldern Brandenburgs. Waldmoorschutzprogramm, Erfolge, Renaturierungsmaßnahmen. Hochschule für
Nachhaltige Entwicklung Eberswalde, Humboldt Universität zu Berlin. Eberswalde, Berlin: 43 S. DOI: 10.57741/opus4-366
↑
Hanspach J., Kühn I., Klotz S. (2013): Risikoabschätzung für Pflanzenarten, Lebensraumtypen und ein funktionelles Merkmal. In: Vohland
K., Badeck F. et al. (Hrsg.): Schutzgebiete Deutschlands im Klimawandel – Risiken und Handlungsoptionen. Naturschutz und Biologische
Vielfalt 129: 71 – 85.
↑
Hauck M., Leuschner C., Homeier J. (2019): Klimawandel und Vegetation. Springer Spektrum. Heidelberg:
358 S.
↑
Hofer B., Köbbing J. (2021): Faktencheck – Was bedeutet die Vorgabe der Deutschen Moorschutzstrategie
„Einsparung von Emissionen in Höhe von 5 Millionen t CO2 Äquivalenten pro Jahr aus Moorböden in Deutschland bis 2030“ in der
praktischen Umsetzung? TELMA 51: 153 – 164.
↑
Jedicke E. (Hrsg.) (1997): Die Roten Listen. Gefährdete Pflanzen, Tiere, Pflan-zengesellschaften und
Biotoptypen in Bund und Ländern. Ulmer Verlag. Stuttgart: 581 S.
↑
Koska I., Succow M., Timmermann T. (2001): Vegetationsformen der offenen, naturnahen Moore und des
aufgelassenen Feuchtgrünlandes. In: Succow M., Joosten H. (Hrsg.): Landschaftsökologische Moorkunde. 2. Aufl. E. Schweizerbart'sche
Verlagsbuchhandlung. Stuttgart: 144 – 161.
↑
Landgraf L. (2007): Zustand und Zukunft der Arm- und Zwischenmoore in Brandenburg: Bewertung und
Bilanz. Naturschutz und Landschaftspflege in Brandenburg – Beiträge zu Ökologie, Natur- und Gewässerschutz 16: 104 – 115.
↑
LUA/Landesamt für Umwelt Brandenburg (Hrsg.) (2004): Leitfaden zur Renaturierung von Feuchtgebieten in Brandenburg. Studien- und Tagungsberichte
50: 88 S.
↑
LUA/Landesumweltamt Brandenburg (Hrsg.) (2006): Liste und Rote Liste der etablierten Gefäßpflanzen Brandenburgs. Naturschutz
und Landschaftspflege in Brandenburg 2006(4): 11 S.
↑
Luthardt V. (2014): Moore im Funktionsgefüge der Landschaft. In: Luthardt V., Zeitz J. (Hrsg.): Moore
in Brandenburg und in Berlin. Natur und Text. Rangsdorf: 14 – 17.
↑
Luthardt V., Brauner O. et al. (2005): Lebensräume im Wandel – Bericht zur ökosystemaren Umweltbeobachtung (ÖUB) in den
Biosphärenreservaten Brandenburgs. Fachbeiträge LUA Brandenburg 94: 188 S.
↑
Luthardt V., Brauner O. et al. (2017): Methodenkatalog zum Monitoring-Programm der Ökosystemaren Umweltbeobachtung in den
Biosphärenreservaten Brandenburgs. 5., akt. Ausg. Teil A: Terrestrische Ökosysteme, Teil B: Aquatische Ökosysteme. Hochschule für
Nachhaltige Entwicklung Eberswalde. Eberswalde: 177 S., 134 S. + Anhänge.
↑
Luthardt V., Brauner O. et al. (2022):
Steckbriefe und weiterführende Informationen zu ausgewählten Mooren des Biosphärenreservats Schofheide-Chorin. Hochschule für
Nachhaltige Entwicklung Eberswalde. Eberswalde: 41 S. DOI: 10.57741/opus4-377
↑
Luthardt V., Klawitter J. (2014): Gefäßpflanzenarten und Moosarten der Moore und Feuchtgebietsarten mit
enger Bindung an Moore in Brandenburg und Berlin. In: Luthardt V., Zeitz J. (Hrsg.): Moore in Brandenburg und in Berlin. Natur und
Text. Rangsdorf: DVD II-3.
↑
Luthardt V., Lüdicke T. et al. (2021): Erfolgreiche Revitalisierung naturnaher Moore im Naturpark
Stechlin-Ruppiner Land. In: Scherfose V. (Hrsg.): Erfolgskontrollen im Naturschutz. BfN-Skripten 171: 169 – 191.
↑
Luthardt V., Wichmann S. (2016): Ecosystem services of peatlands. In: Wichtmann W., Schröder C.,
Joosten H. (Hrsg.): Paludiculture – Productive use of wet peatlands: Climate protection – biodiversity – regional economic benefits.
Schweizerbart Science Publishers. Stuttgart: 13 – 20.
↑
Luthardt V., Zeitz J. (Hrsg.) (2014): Moore in Brandenburg und in Berlin. Natur und Text. Rangsdorf:
384 S.
↑
Metzing D., Hofbauer N. et al. (Red.) (2018): Rote Liste gefährdeter Tiere, Pflanzen und Pilze
Deutschlands. Pflanzen. Naturschutz und Biologische Vielfalt 70(7): 784 S.
↑
MLUL Brandenburg/Ministerium für Ländliche Entwicklung, Umwelt und Landwirtschaft des Landes Brandenburg (Hrsg.) (2016): Moorschutz im brandenburgischen Wald. 3. Aufl. MLUL. Potsdam: ohne Seitenzahl.
↑
Petermann J., Balzer S. et al. (2007): Klimawandel – Herausforderungen für das europaweite
Schutzgebietssystem Natura 2000. In: Balzer S., Dietrich M., Beilich B. (Hrsg.): Natura 2000 und Klimaänderungen. Naturschutz und
Biologische Vielfalt 46: 127 – 148.
↑
Pompe S., Berger S. et al. (2011): Modellierung der Auswirkungen des Klimawandels auf Flora und
Vegetation in Deutschland. BfN-Skripten 304: 193 S.
↑
Reichelt K. (2015): Evaluierung des GEST-Modells zur Abschätzung der Treibhausgasemissionen aus Mooren.
Masterarbeit Universität Greifswald. Greifswald: 39 S.
↑
Roth M., Succow M. (2001): Vegetationsformen des Saatgraslandes, des Intensivgraslandes und daraus
wieder hervorgehender Wiesen und Weiden. In: Succow M., Joosten H. (Hrsg.): Landschaftsökologische Moorkunde. 2. Aufl. E.
Schweizerbart'sche Verlagsbuchhandlung. Stuttgart: 171 – 181.
↑
SCB/Society for Conservation Biology (2021): International Congress for Conservation Biology (ICCB).
Internetartikel. https://conbio.org/conferences/about-scb-meetings/ (aufgerufen am 1.12.2022).
↑
Spangenberg A. (2011): Einschätzung der Treibhausgasrelevanz bewaldeter Moorstandorte in Mecklenburg-Vorpommern
hinsichtlich des Minderungspotentials nach Wiedervernässung. Endbericht für DUENE e. V. Greifswald: 29 S.
↑
Succow M. (2001): Übersicht chorischer Moor-Naturraumtypen. In: Succow M., Joosten H. (Hrsg.):
Landschaftsökologische Moorkunde. Schweizerbart'sche Verlagsbuchhandlung. Stuttgart: 240 – 252.
↑
Succow M., Joosten H. (Hrsg.) (2001): Landschaftsökologische Moorkunde. Schweizerbart'sche
Verlagsbuchhandlung. Stuttgart: 622 S.
↑
Tanneberger F., Appulo L. et al. (2021): The power of nature-based solutions: How peatlands can help us to achieve key EU sustainability objectives. Advanced
Sustainable Systems 5(1): 2000146. DOI: 10.1002/adsu.202000146
↑
Tiemeyer B., Borraz E. et al. (2020): A new methodology for organic soils in national greenhouse gas inventories: Data synthesis, derivation and
application. Ecological Indicators 109: 105838. DOI: 10.1016/j.ecolind.2019.105838
↑
Trepel M., Pfadenhauer J., Zeitz J. et al. (2017): Germany. In: Joosten H., Tanneberger F.,
Moen A. (Hrsg.): Mires and peatlands of Europe – Status, distribution and conservation. Schweizerbart Science Publishers. Stuttgart:
413 – 425.
↑
UBA/Umweltbundesamt (Hrsg.) (2022): Emissionen der Landnutzung, -änderung und Forstwirtschaft. https://bit.ly/UBA-LULUCF (aufgerufen am 1.12.2022).
Zusatzmaterial zum Beitrag
Zusatzmaterial (1 Erfassungsjahre und
Vegetationsparameter der naturnahen Moore innerhalb der Ökosystemaren Umweltbeobachtung, 2 Wasserstufen und Wasserstufenausbildungen
für Moore und andere Feuchtgebiete, 3 Das Indikatorensystem zur Bewertung moorspezifischer Biodiversität, 4 Mittlere jährliche Dauer
der Vegetationsperiode an den Meldestationen im Biosphärenreservat Schorfheide-Chorin, 5 Übersicht der in den naturnahen Mooren
erfassten moorspezifischen 6 Gesamtartenzahl der Gefäßpflanzen und Moose und Artenzahl der Teilmenge der moorspezifischen Arten in den
Vegetationsaufnahmen Gefäßpflanzen und Moose mit Angaben zur Gefährdung, 7 Detaillierte Ergebnisse des Indexes der moorspezifischen
Biodiversität, 8 Literatur) als PDF herunterladen (PDF 1.254 kB)

