Silke Lütt
Zusammenfassung
Auf 22 Dauerbeobachtungsflächen in drei Mooren Schleswig-Holsteins wurde von 1989 bis 2019 im Abstand von 10 Jahren die Vegetation erfasst. In zahlreichen Handtorfstichen konnten die Ausbreitung von Mittlerem Torfmoos (Sphagnum magellanicum) und die Entwicklung der
Hochmoorbultengesellschaft über eine Zeitspanne von 30 Jahren sowie eine Oligo- und Ombrotrophierung (Verringerung der Nähstoffversorgung und Ernährung aus dem Niederschlagswasser) der Standorte beobachtet werden. Stoppen der Binnenentwässerung, Überstau, Abschieben und
Birkenentnahmen haben zur Revitalisierung schützenswerter Lebensräume geführt. Im Fockbeker Moor wurden beim Erico-Sphagnetum typicum in den letzten 10 Jahren eine Abnahme der Schlenken (tiefer gelegene Standorte im Mikrorelief von Mooren), eine stärkere Bildung von Bulten
(höhere Standorte) und eine Zunahme der Heidekräuter festgestellt. Auf einer nackten Torffläche sind nach 23 Jahren Torfmoose eingewandert. Die Sukzession der überstauten Fläche im Fockbeker Moor begann mit Torfmoosrasen, Eriophorum-Arten und Heidekräutern nach
10 Jahren und zeigt heute Bewaldung mit Moor-Birke (Betula pubescens). In fast allen Dauerflächen ist eine Zunahme der Phanerogamendeckung und der Artenzahlen festzustellen. Seit 10 Jahren ist eine Ausbreitung von Weißem Schnabelried (Rhynchospora alba) und
Besenheide (Calluna vulgaris) im Wittenseer und Fockbeker Moor und von Moorlilie (Narthecium ossifragum) im Owschlager Moor zu verzeichnen. Die Veränderung der Artenzusammensetzung und die Ombrotrophierung werden dargestellt und vor dem Hintergrund durchgeführter
Revitalisierungsmaßnahmen, autogener Sukzession, Stickstoffbelastung und Klimawandel interpretiert. Es werden Empfehlungen für künftig stärker zu berücksichtigende Maßnahmen in Hinblick auf den Klimawandel gegeben.
Sukzession – Moor – Stickstoffdeposition – Klimawandel – Erico-Sphagnetum magellanici – Sphagnum – RevitalisierungsmaßnahmenAbstract
On 22 permanent plots in three mires, vegetation development was investigated at intervals of 10 years over a period of 30 years. Most of the plots in peat cuttings exhibited a spreading of Sphagnum magellanicum and a development of Erico-Sphagnetum
magellanici within 10 to 30 years, indicating oligo- and ombrotrophication of the sites. Ceasing dehydration, ensuring overflow, stripping toppeat and removing birches are actions that have revitalised rare and protected bog sites. The plots with Erico-Sphagnetum typicum
in Fockbeker Moor exhibit an increasing microtopography combined with shrub enrichment within the last 10 years. On nude peat areas, recovery of Sphagnum species takes 23 years. Succession on large rewetted pools in Fockbeker Moor started with Sphagnum
layers, Eriophorum and ericoid species after 10 years and exhibits stands of Betula pubescens today. Nearly all plots indicate an increasing species richness and greater area coverage by vascular plants. Since 10 years, expansion of Rhynchospora alba
and Calluna vulgaris in Wittenseer und Fockbeker Moor and of Narthecium ossifragum in Owschlager Moor can be found. The present article interprets changes in species composition and ombrotrophication with regard to revitalisation management, autogenic succession,
nitrogen deposition and climate change. It makes recommendations for restoration measures that will need to gain greater priority under climate change.
Succession – Mire – Nitrogen deposition – Climate change – Erico-Sphagnetum magellanici – Sphagnum – Restoration measuresInhalt
1 Einleitung
Aktuell hat die Wiedervernässung von Mooren durch den Klimaschutz eine neue Bedeutung gewonnen. In Schleswig-Holstein werden jedoch schon seit mehr als 60 Jahren Hochmoorreste renaturiert (LLUR 2015). Entscheidendes Etappenziel bei
dieser Renaturierung ist die Wiederausbreitung oder Neuansiedlung des Torfbildners Sphagnum magellanicum (zu den deutschen Artnamen aller hier im Beitrag genannten Pflanzenarten siehe Tab. B, C, D im Online-Zusatzmaterial II). Zielvegetation ist die Hochmoorbultengesellschaft (Bulte = etwas erhöhte Standorte im Mikrorelief von Mooren).
Doch ist dieses Ziel bei zunehmenden Stickstoffeinträgen und fortschreitendem Klimawandel noch zu erreichen? Vor diesem Hintergrund kommt Langzeituntersuchungen zur Vegetationsentwicklung eine besondere Bedeutung zu. Sie helfen, Möglichkeiten und Grenzen der Restitution von
Hochmoorresten in Zeiten des globalen Wandels zu bewerten und Ziele des Moorschutzes zu überprüfen.
Seit 1881 ist für Schleswig-Holstein ein kontinuierlicher Anstieg der Jahresmitteltemperatur um 1,3 °C (DWD 2017) belegt. Gleichzeitig haben die Niederschläge im gleichen Zeitraum um 18 % auf durchschnittlich 823 mm
zugenommen (auch Starkregenereignisse, DWD 2017). Die ganzjährig steigende Temperatur schlägt sich in erhöhten Evapotranspirationsraten (Summe aus Evaporation und Transpiration, d. h. Verdunstung von Wasser von Boden- und
Wasseroberflächen sowie aus Pflanzen und Tieren) nieder, die immer weniger durch den für das atlantische Klima Schleswig-Holsteins typischen gemäßigten Sommerregen kompensiert werden. Langfristige Prognosen bestätigen diesen Trend (DWD 2017). Eine
Zunahme von Grasartigen und Bäumen als Folge des Klimawandels wurde für die Moore Nordeuropas bereits nachgewiesen (Breeuwer et al. 2009; Heijmans et al. 2013).
Kurzfristig folgenschwerer als der Klimawandel wirkt sich für Hochmoore die Stickstoffbelastung aus, die zu Veränderungen des Artengefüges führt (Twenhöven 1992; Wiedermann et al. 2007;
Bobbink et al. 2011). Feld- und Laborexperimente lassen erwarten, dass die Etablierung der Zielart Sphagnum magellanicum oder gar die Entwicklung von Hochmoorbultengesellschaften durch Veränderungen des Konkurrenzverhaltens
zunehmend schwieriger wird.
Vor 30 Jahren wurden Dauerbeobachtungsflächen angelegt. Erste Ergebnisse dazu wurden nach zehn Jahren publiziert (Lütt 2001). Die hier vorgestellten Ergebnisse fassen die Entwicklung der Vegetation in der Zeitspanne von 1989
bis 2019 anhand prägnanter Fallbeispiele zusammen. Naturschutzfachliches Ziel dabei ist, die Entwicklung torfbildender Vegetation vor dem Hintergrund durchgeführter und zukünftiger Renaturierungsmaßnahmen zu bewerten. Eine ausführliche vegetationskundliche Beschreibung der
Entwicklung sämtlicher Dauerbeobachtungsflächen ist bei Lütt 2020 nachzulesen, ergänzende Ergebnisse finden sich im Online-Zusatzmaterial I sowie in Tab. B, C, D im Online-Zusatzmaterial II.
2 Methoden zur Aufnahme der Vegetation
Im Jahr 1989 wurden in drei Hochmoorresten Schleswig-Holsteins im Kreis Rendsburg-Eckernförde 22 Dauerquadrate (bezeichnet mit D1, D2, U1 etc.) angelegt. Die Größe der Flächen variiert zwischen 1 und 4 m2. Die Flächen wurden markiert und
später mit GPS eingemessen und detailliert fotografiert. Auf den Dauerquadraten wurde die Vegetation der Gefäßpflanzen und Moose aufgenommen. Die Schätzung der Deckungen erfolgte nach der Skala von Londo (1975). Für die Darstellung der Ergebnisse
in Graphiken wurden die mittleren Deckungswerte der Klassen nach Londo in den Aufnahmen von 1989 mit jenen von 2019 verglichen und als prozentuale Zu- oder Abnahme dargestellt. Die Nomenklatur der Gefäßpflanzen folgt Wisskirchen, Haeupler
(1998), jene der Moose Sauer, Ahrends (2006), die der Pflanzengesellschaften Dierßen (1988).
3 Die Untersuchungsgebiete (siehe Abb. 1)
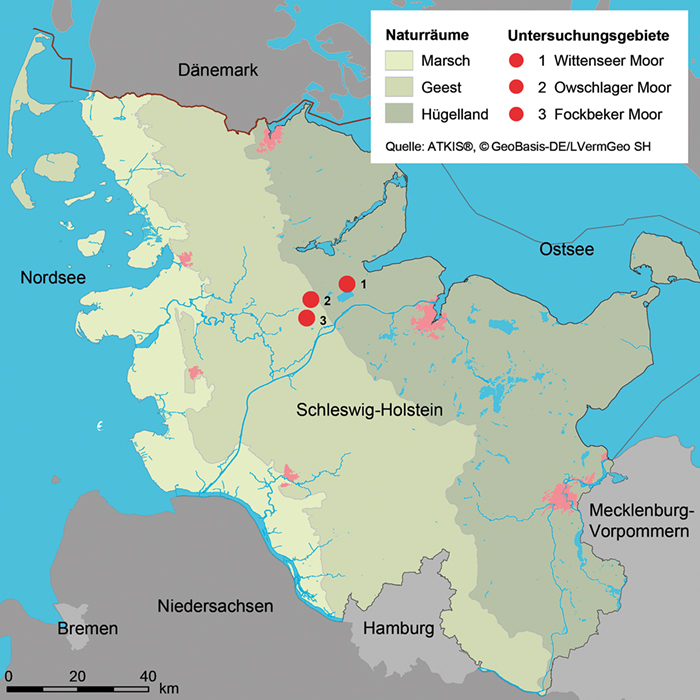
Abb. 1: Lage der untersuchten Moore in Schleswig-Holstein.
Fig. 1: Location of the mires investigated in Schleswig-Holstein.
Das Groß Wittenseer Moor ist ein kleines Hochmoor der schleswig-holsteinischen Jungmoräne am Rande der Hüttener Berge mit einer Größe von nur 14,7 ha. Im baumfreien Moorkörper dominierten 1989 oligotraphente Verlandungsgesellschaften (Gesellschaften an Standorten mit
geringer Nährstoffversorgung) und Initialphasen der Hochmoorgesellschaft.
Das Owschlager Moor liegt im Naturraum der schleswig-holsteinischen Vorgeest. Die Dauerbeobachtungsflächen befinden sich in einem moorgenetisch eigenständigen Moorkörper der durch den Zustrom quellig austretenden Druckwassers eine abweichende Entwicklung nahm. In dem nur
wenige Hektar großen Moorteil kamen 1989 in jüngeren Torfstichen minerotraphente Verlandungsstadien (Stadien an Standorten mit mäßiger Nährstoffversorgung) und Initialen der Hochmoorbultengesellschaft vor.
Das 190 ha große Fockbeker Moor ist ein ursprünglich ausgedehntes, wurzelechtes Hochmoor, das Anfang der 1980er-Jahre großflächig bis über einen Meter überstaut wurde (siehe Abb. 2; Brehm,
Pfeiffer 1982). Die Flächen wurden 1989 von Eriophorum angustifolium-Sphagnum fallax-Schwingdecken eingenommen. Ostwärts schließt eine ausgedehnte Zone kleiner Handtorfstiche mit oligotraphenten Verlandungsgesellschaften bis hin zu Erico-Sphagneten an.

Abb. 2: Anstaufläche im Fockbeker Moor mit oligotraphenten Verlandungsgesellschaften (Gesellschaften an Standorten mit geringer Nährstoffversorgung) Anfang der 1990er-Jahre.
Fig. 2: Huge rewetted pools in Fockbeker Moor with oligotrophic vegetation recolonising in the early 1990s.
(Foto: Kuno Brehm)
Bei sämtlichen Moorresten wurde zum Zweck der Renaturierung die Binnenentwässerung gestoppt. Im Fockbeker Moor erfolgte zusätzlich eine wiederkehrende Birkenentnahme (Entkusselung) sowie das Abschieben mineralisierter Torfschichten und Einsäen von Calluna vulgaris (siehe
Tab. 1).
|
Tab. 1: Übersicht zu Renaturierungsmaßnahmen und Vegetationsveränderungen in den Dauerflächen im Fockbeker, Wittenseer und Owschlager Moor (Schleswig-Holstein) im Verlauf von 30 Jahren (1989 – 2019).
Table 1: Overview of restoration measures and vegetation succession in permanent plots in Fockbeker Moor, Wittenseer Moor und Owschlager Moor (Schleswig-Holstein) over 30 years (1989 – 2019).
|
|
Maßnahme
|
Dauerfläche
|
Vegetation 1989
|
Vegetation 2019
|
|
Fockbeker Moor
|
|
Überstau 1982
|
D4, U1
|
Eriophorum angustifolium-Gesellschaft (Gesellschaft des Schmalblättrigen Wollgrases), Fazies von Sphagnum fallax
|
Eriophorum angustifolium-Gesellschaft (Gesellschaft des Schmalblättrigen Wollgrases) in Entwicklung zum Betuletum pubescentis (Moorbirkenwald, D4) oder Erico-Sphagnetum magellanici (Hochmoorbultengesellschaft, U1)
|
|
Abschieben mineralisierter Weißtorfe; Einsaat von Calluna vulgaris
|
D8
|
Eriophorum vaginatum-Gesellschaft (Gesellschaft des Scheidigen Wollgrases), Fazies von Dicranella cerviculata und Campylopus pyriformis
|
Erico-Sphagnetum magellanici sphagnetosum cuspidati, Phase von Rhynchospora alba (Hochmoorbultengesellschaft in der Subassoziation von Sphagnum cuspidatum in der Phase des Weißen Schnabelrieds)
|
|
Stoppen der Binnenentwässerung
|
D6
|
Erico-Sphagnetum magellanici typicum (Hochmoorbultengesellschaft)
|
Erico-Sphagnetum magellanici, Phase von Calluna vulgaris (Hochmoorbultengesellschaft in der Phase von Calluna vulgaris)
|
|
Wittenseer Moor
|
|
Stoppen der Binnenentwässerung
|
D4, D7
|
Eriophorum angustifolium-Gesellschaft (Gesellschaft des Schmalblättrigen Wollgrases)
|
Erico-Sphagnetum magellanici, Phase von Rhynchopora alba (Hochmoorbultengesellschaft, Phase des Weißen Schnabelrieds)
|
|
Owschlager Moor
|
|
Stoppen der Binnenentwässerung
|
D2
|
Eriophorum angustifolium-Gesellschaft (Gesellschaft des Schmalblättrigen Wollgrases)/Scorpidio-Utricularietum minoris (Gesellschaft des Kleinen Wasserschlauchs)
|
Erico-Sphagnetum magellanici, Subassoziation von Narthecium ossifragum (Hochmoorbultengesellschaft in der Subassoziation von Narthecium ossifragum)
|
|
Subassoziation: Pflanzengesellschaft unterhalb der Rangstufe einer Assoziation, die in einem größeren Teilgebiet der Verbreitung der Assoziation (Grundeinheit der Pflanzensoziologie) vorkommt.
Fazies: Aspekt einer Pflanzengesellschaft, der durch das Vorherrschen einer oder weniger Arten geprägt wird.
Phase: Abschnitt im Verlauf einer Sukzession mit Auftreten besonderer diesen Abschnitt kennzeichnender Arten (nach Dierschke 1994).
|
4 Die Vegetationsentwicklung in den Dauerquadraten von 1989 bis 2019 [1]
4.1 Veränderung der Deckungen von Phanerogamen und Kryptogamen sowie der Artenzahl (siehe Abb. 3)
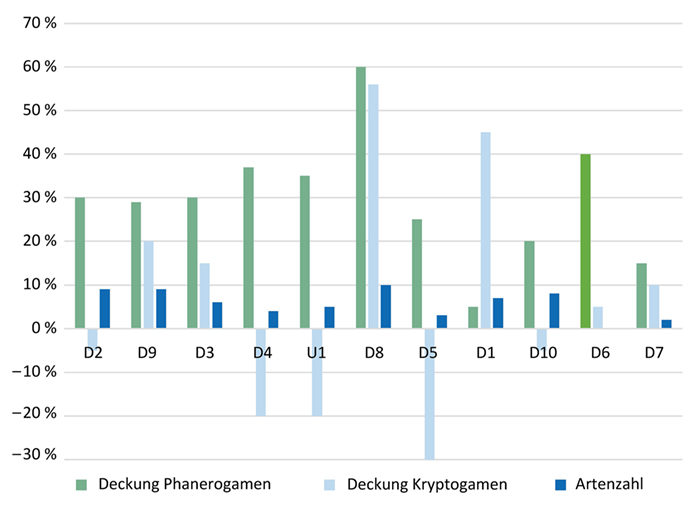
Abb. 3: Zu- und Abnahmen der prozentualen Deckungen der Phanerogamen und Kryptogamen sowie der Artenzahlen exemplarisch dargestellt für die Dauerflächen des Fockbeker Moors über die Zeitspanne 1989 – 2019.
Fig. 3: Increase and decrease of percentage cover of plants, bryophytes and number of species, exemplified by permanent plots in Fockbeker Moor over the period from 1989 to 2019.
Im Vergleich zur mittleren Deckung der Phanerogamen im Jahr 1989 ist jene im Jahr 2019 in allen Dauerquadraten in allen drei Mooren angestiegen, der Median der prozentualen Zunahme liegt im Fockbeker Moor bei 30 %, im Wittenseer Moor bei 12 % und im
Owschlager Moor bei 48 %. Die Deckung der Kryptogamen, hier insbesondere der Torfmoose, ist in einigen Dauerflächen angestiegen, in anderen gesunken. Die Artenzahl ist, mit Ausnahme der Hochmoorbultengesellschaft (D6), um 1 – 9 Arten angestiegen (Lütt 2020, siehe auch Abb. I, L im Online-Zusatzmaterial I). Die Zuordnung der Dauerquadrate zu den
Maßnahmen ist in Tabelle 1 dokumentiert.
4.2 Fockbeker Moor
Ausgangsgesellschaft Eriophorum angustifolium-Gesellschaft Fazies von Sphagnum fallax (U1, D4)
Die beiden Dauerquadrate befinden sich in der großflächigen Überstaufläche (D4, U1). Die Flächen nehmen eine unterschiedliche Entwicklung: Während in D4 Eriophorum vaginatum, Molinia caerulea, Calluna vulgaris und Betula pubescens zunehmen (siehe Abb. 4) und eine Entwicklung zum Moorbirkenwald anzeigen (siehe auch D1, D5, Abb. E, F im Online-Zusatzmaterial I), weist U1 sowohl Kennarten der Hochmoorbultengesellschaft als auch des Moorbirkenwalds auf und befindet sich in Hinblick auf die Bestandsentwicklung in einer Interimsphase (siehe Abb. 5).
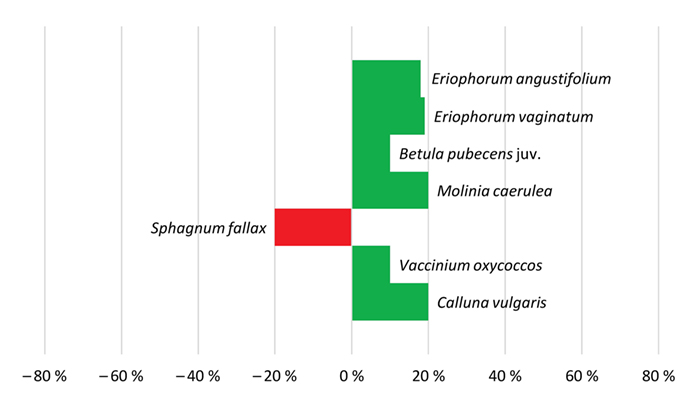
Abb. 4: Zu- und Abnahmen der prozentualen Deckungen von Arten in der Dauerfläche D4 im Fockbeker Moor über die Zeitspanne 1989 – 2019.
Fig. 4: Increase and decrease of percentage cover of species in permanent plot D4 in Fockbeker Moor over the period from 1989 to 2019.
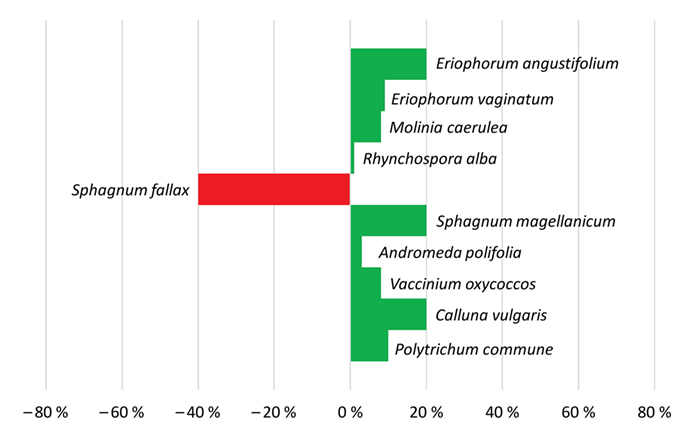
Abb. 5: Zu- und Abnahmen der prozentualen Deckungen von Arten in der Dauerfläche U1 im Fockbeker Moor über die Zeitspanne 1989 – 2019.
Fig. 5: Increase and decrease of percentage cover of species in permanent plot U1 in Fockbeker Moor over the period from 1989 to 2019.
Mit zunehmendem Alter vergreisen die Bulte von Eriophorum vaginatum und werden bereits nach zehn Jahren von Laub- und Lebermoosen, Torfmoosen und auch Gefäßpflanzen besiedelt. Die Wirkung der Bulten als „Ammen“ wurde auch von Sliva
(1997), Daniels (2001), Nick, Weber (2001) beschrieben. Die Genese von Moorbirkenwäldern im Zuge einer Hochmoorbildung wurde für viele Moore Schleswig-Holsteins anhand von Torfprofilen belegt
(Wagner 1994). Die hohen Stickstofffrachten begünstigen außerdem die Bewaldung (Hejmanns et al. 2013). Auch für Eriophorum vaginatum oder Molinia caerulea, die sich in einigen
Dauerquadraten des Fockbeker Moores ausbreiten konnten (z. B. D4, U1), wurde eine Förderung durch Stickstofffrachten nachgewiesen (Aaby 1994; Schmidt 2014).
Ausgangsgesellschaft Eriophorum vaginatum-Gesellschaft, Fazies von Dicranella cerviculata und Campylopus pyriformis (D8)
Dieser trockenste Flügel der Gesellschaft besiedelte im Jahr 1986 freigeschobene Weißtorfe im Zentrum des Fockbeker Moores. Der Bereich selbst ist nicht angestaut, profitiert aber durch einen Rückstau vom Überstau. In den letzten 20 Jahren sind die Flächen
kontinuierlich nasser geworden. Die im Jahr 1989 nackte Fläche entwickelte sich über eine Phase mit wenigen Arten, dominiert von Eriophorum vaginatum, Calluna vulgaris und Pionierarten wie Drosera rotundifolia sowie den Laubmoosen, seit 1999 allmählich in
Richtung Hochmoorbultengesellschaft. Das Einsäen von Calluna vulgaris im Jahr 1986 zeigte zunächst keinen Effekt, erst 1999 stieg die Deckung bis auf 50 %. Torfmoose haben sich erst 2009 mit Sphagnum cuspidatum, S. rubellum und
S. papillosum eingestellt. Die Zunahme von Rhynchospora alba, Hypnum jutlandicum und Jungbirken zeigt die Wechselfeuchte des Standorts an (Abb. 6).
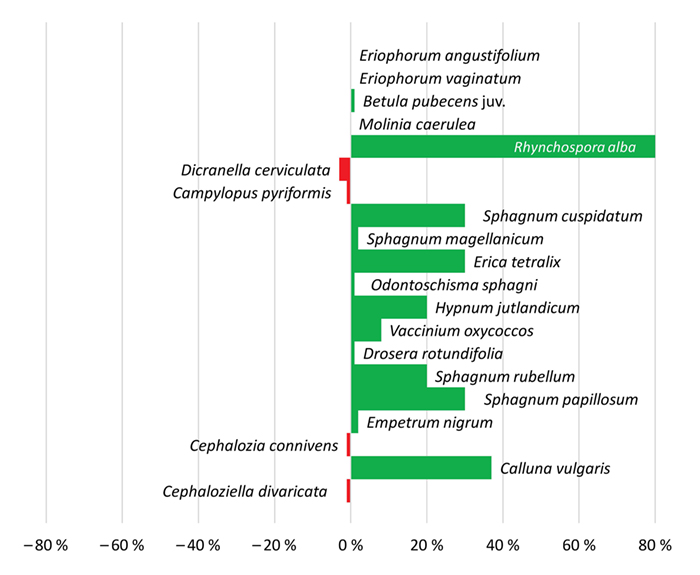
Abb. 6: Zu- und Abnahmen der prozentualen Deckungen von Arten in der Dauerfläche D8 im Fockbeker Moor über die Zeitspanne 1989 – 2019. Ohne Balken bedeutet unverändert.
Fig. 6: Increase and decrease of percentage cover of species in permanent plot D8 in Fockbeker Moor over the period from 1989 to 2019. Without bar means no change.
Nackte Torfflächen sind langwährende Stadien nach industriellem Torfabbau. Erst nach mehr als 20 Jahren haben sich auf der abgeschobenen Weißtorffläche Torfmoose und Hochmoorarten angesiedelt. Der dunkle Torf ist mikroklimatisch und physikalisch ein schwieriger Lebensraum mit
Kammeisbildung im Winter, extrem hohen Temperaturen im Sommer, Nährstoffarmut sowie fehlenden Mykorrhizapilzen (Poschlod 1990). Eriophorum vaginatum ist hier ein typischer Primärbesiedler (Sliva
1997). Durch die Wassersättigung infolge des Überstaus haben sich Wasserstände und Trophie (Nährstoffangebot eines Standorts) auf ein Schlenkenniveau (Schlenken = etwas tiefer gelegene Standorte im Mikrorelief von Mooren) eingestellt und eine mykologische
Reifung hat die Fläche erst nach einem Jahrzehnt für Calluna vulgaris und Torfmoose bewohnbar gemacht. Samen von Calluna vulgaris sind langlebig (Poschlod 1991; Waesch 2006), reagieren aber
u. a. empfindlich auf Trockenheit (Härdtle et al. 2009).
Ausgangsgesellschaft Erico-Sphagnetum magellanici typicum (D6)
Die deutlichste Veränderung in den ersten zehn Jahren ist die Zunahme von Erica tetralix, Andromeda polifolia und von Sphagnum magellanicum. Die S. cuspidatum-Schlenke war stabil. Zwanzig Jahre später hat sich das Mikrorelief zugunsten der Bulte
verschoben, der Deckungsanteil der Gefäßpflanzen insgesamt erhöht und jener der Kryptogamen verringert. Calluna vulgaris hat mit 40 % Deckung deutlich zugenommen, ebenso Eriophorum vaginatum und Vaccinium oxycoccos. Die Schlenkenbewohner
S. cuspidatum, S. fallax, S. papillosum, E. angustifolium sowie das Lebermoos Cephalozia connivens weisen entsprechend niedrigere Deckungsanteile auf. S. magellanicum ist mit gleichbleibend hoher Deckung
vertreten und vital. Auf den Bulten beginnt sich Hypnum jutlandicum auszubreiten (siehe Abb. 7).
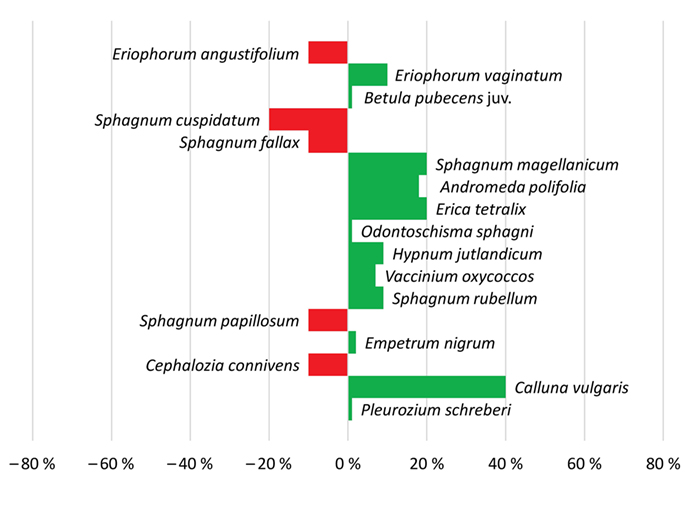
Abb. 7: Zu- und Abnahmen der prozentualen Deckungen von Arten in der Dauerfläche D6 im Fockbeker Moor über die Zeitspanne 1989 – 2019.
Fig. 7: Increase and decrease of percentage cover of species in permanent plot D6 in Fockbeker Moor over the period from 1989 to 2019.
Der Bestand der Hochmoorbultengesellschaft in D6 (Abb. 7) zeigt nach 30 Jahren eine Zunahme der Bulten mit Calluna vulgaris und eine Abnahme der Schlenkenarten. Dies kann ein Effekt stärker schwankender Wasserstände und
trockenerer Sommermonate der letzten Jahre sein. Ein ausgeprägtes Mosaik von Bulten und Schlenken ist aber auch typisch für kontinentale Moorlandschaften mit zeitweiligem Niederschlagsdefizit (Dierßen 2006; Kollmann
2019). Auch die ehemalige Mooroberfläche in D7 (Abb. G im Online-Zusatzmaterial I) zeigt vergleichbare Trockenheitseffekte.
4.3 Wittenseer Moor
Ausgangsgesellschaft Eriophorum angustifolium-Gesellschaft (D4, Abb. 8, D7)
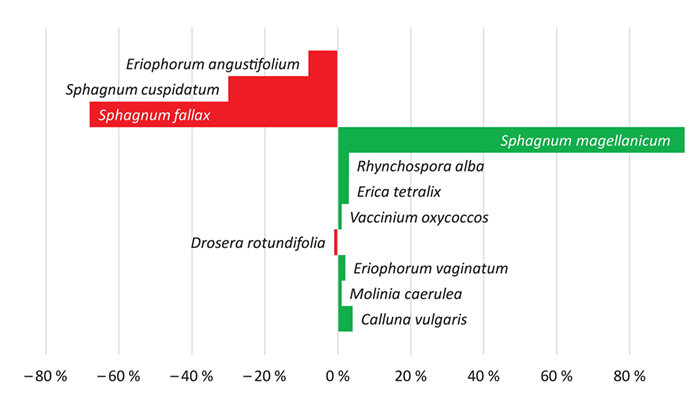
Abb. 8: Zu- und Abnahmen der prozentualen Deckungen von Arten in der Dauerfläche D4 im Wittenseer Moor über die Zeitspanne 1989 – 2019.
Fig. 8: Increase and decrease of percentage cover of species in permanent plot D4 in Wittenseer Moor over the period from 1989 to 2019.
Sphagnum fallax war 1989 das dominierende Torfmoos. Zwanzig Jahre später ist S. magellanicum mit einer Deckung von 95 % vorherrschend auf dem Rasen. Sowohl S. fallax als auch S. cuspidatum sind ebenso wie Eriophorum
angustifolium deutlich zurückgegangen. Molinia caerulea hatte zwischenzeitlich zugenommen, ging in den Folgejahren allerdings in den Deckungsanteilen zurück. Mit der Ansiedlung von Calluna vulgaris und vereinzelt auch E. vaginatum zwanzig Jahre später beginnt
der Rasen 2019 zu flachen Bulten aufzuwachsen, insgesamt ist der Anteil an Gefäßpflanzen mit etwa 20 % noch ausgesprochen niedrig.
Auch in den nassen Stadien der Hochmoorbultengesellschaft entwickelten sich über die betrachtete Zeitspanne von 30 Jahren die Watten und Teppiche zu Rasen mit Deckungsanteilen von Sphagnum magellanicum von mehr als 85 % im Jahr 2019. Schäden durch
Wildtritt waren bereits nach zehn Jahren komplett wieder von S. magellanicum und S. cuspidatum geschlossen (siehe Abb. J, K im Online-Zusatzmaterial I, Tab. C im Online-Zusatzmaterial II). Die auffälligste Veränderung seit 2009 ist die
Ausbreitung von Rhynchospora alba.
4.4 Owschlager Moor
Ausgangsgesellschaft Eriophorum angustifolium-Gesellschaft (D2, Abb. 9)

Abb. 9: Zu- und Abnahmen der prozentualen Deckungen von Arten in der Dauerfläche D2 im Owschlager Moor über die Zeitspanne 1989 – 2019.
Fig. 9: Increase and decrease of percentage cover of species in permanent plot D2 in Owschlager Moor over the period from 1989 to 2019.
Bereits nach zehn Jahren hatten die Schwingdecken von Sphagnum fallax und S. papillosum den Wasserkörper mit der Wasserschlauch-Gesellschaft und Watten von S. denticulatum mehr oder weniger verdrängt und die Deckungen der Niedermoorarten gingen
zurück. Bis 2009 setzte sich dieser Prozess fort, Narthecium ossifragum und S. magellanicum siedelten sich zu einem dichten, massiven Rasen an. Weitere zehn Jahre später sind die Deckungen der Hochmoorarten nochmals angestiegen, insbesondere durch Narthecium
ossifragum (siehe Abb. M, N im Online-Zusatzmaterial I, Tab. D im Online-Zusatzmaterial II).
4.5 Naturschutzmaßnahmen
Hervorzuhebender Erfolg in sämtlichen Mooren ist die Ausbreitung von Sphagnum magellanicum (Rote-Liste-Kategorie 3 „Gefährdet“, Caspari et al. 2018) und die Entwicklung der stark gefährdeten Hochmoorbultengesellschaft
(Rennwald 2000) in den Torfstichen, wodurch die selbstregulierenden Fähigkeiten dieser Moorreste (Lütt 1992; Succow, Joosten 2001; Timmermann et al. 2009) mit S. magellanicum als „ecosystem engineer“ (van Breemen 1995; Hölzel et al. 2019; Raabe et al. 2018) wiederhergestellt werden. Torfstiche sind damit wichtige Ausgangspunkte für die Wiederbesiedlung der umgebenden Flächen. Diese wurde im Wittenseer und Owschlager Moor allein durch das Stoppen der Binnenentwässerung eingeleitet. Als bundesweit
bedroht eingestufte (Finck et al. 2017) Lebensräume einer hochspezialisierten Flora und Fauna wurden hier neu geschaffen, die Kohlenstoff dauerhaft im Torf speichern und außerdem positiv auf den Wasserhaushalt der umgebenden Landschaft
wirken (Naturkapital Deutschland 2014; Kollmann 2019). Auch für renaturierte Hochmoore Mecklenburg-Vorpommerns ist Torfstichregeneration typisch (Gelbrecht et al.
2011) und für viele Moore Süddeutschlands hat das Unterlassen der Grabenpflege sowie das Stoppen der Binnenentwässerung zu günstigen Entwicklungen beigetragen (Kaule, Peringer 2015). In den hier untersuchten Mooren sind
nach 30 Jahren kaum noch Niveauunterschiede zwischen Torfstich und Rändern vorhanden.
Eine zukünftige Transgression der wüchsigen Moorvegetation über die Torfstiche hinaus ist aber nur bei weitreichenderen Anhebungen des Wasserstands der umgebenden Resttorfköper mit Anlage einer hydrologischen Schutzzone möglich (Pfadenhauer, Grootjans
1999; Edom 2001; Bönsel 2011). Erst dann kann der Lebensraumtyp (LRT) 7110 „Lebende Hochmoore“ eine weiträumige Ausbreitung auf Kosten des LRT 7120 „Renaturierungsfähige
degradierte Hochmoore“ erfahren (Ssymank et al. 2015, siehe Kasten 1). Weitere mögliche Entwicklungen – z. B. durch flächigen Überstau – können zu Moorbirkenwäldern
(LRT 91D0*) und sekundären Schlenken (LRT 7150) mit kleinen Hochmoorinitialen führen, wie die Ergebnisse aus dem Fockbeker Moor zeigen. Primäre Moorbirkenwälder sind zudem ein prioritärer Fauna-Flora-Habitat(FFH)-LRT (91D0*, Ssymank
et al. 1998), der bundesweit stark gefährdet ist (Rote-Liste-Kategorie 1 – 2 „Vom Aussterben bedroht“ – „Stark gefährdet“, Fink et al 2017). Sie können sich insbesondere bei zunehmenden
Trockenphasen begünstigend auf das Wachstum der Torfmoose auswirken (Edom 2001; Laube 2009; Bretschneider 2012; Müller-Kröhling
et al. 2019). Lichte Moorwälder sind zudem im Klimawandel wichtige Refugialstandorte für viele Insekten des Hochmoors (Kaule et al. 2018).
Kasten 1: Empfehlungen für Renaturierungsmaßnahmen in Hochmoorresten bei steigenden Klimabelastungen.
Box 1: Recommended restoration measures in raised bogs under increasing climatic stress.
● Stoppen der Binnenentwässerung ist eine wichtige Initialmaßnahme zur Förderung einer hochmoortypischen Vegetation in Torfstichen (siehe auch Bönsel 2011; Kaule, Peringer
2015). ● Kleinflächige bäuerliche Torfstiche dienen als wichtige Wiederausbreitungsherde einer hochmoortypischen Vegetation (siehe auch Lütt 1992; Timmermann et al. 2009), denn die
Ausbreitung und die Samenbank der meisten Moorarten ist nur schwach entwickelt (siehe Kollmann 2019). ● Fehlen diese nach industriellem Torfabbau, können Bunkerde (oberste Vegetationsschicht, die beim Torfabbau abgeräumt wurde), Ammenpflanzen und Impfungen mit Torfmoosen die Wiederausbreitung einer hochmoortypischen Vegetation beschleunigen (Sliva 1997; Hölzel et al. 2019; Raabe et al. 2018).
● Für die Ausbreitung der Hochmoorvegetation aus Torfstichen in die Umgebung ist eine Durchnässung der umgebenden Torfkörper notwendig. Überstau kann helfen die Wassersättigung der Resttorfkörper herbeizuführen (Timmermann et al. 2009), führt jedoch zumindest
zeitweilig zu Methanfreisetzungen (Höper, zitiert in Belting, Blankenburg 2014). ● Oberflächennahe hohe Wasserstände, wie sie sich in Torfstichen aufgrund des Wasserzustroms von den Resttorfkörpern einstellen, sind besonders wichtig. Für Renaturierungsflächen werden mittlere Wasserstände von 10 cm unter Flur empfohlen (Höper in Belting, Blankenburg 2014).
● Geringere Sommerniederschläge, steigende Niederschläge im Winter sowie Starkregenereignisse erfordern zunehmend variable Staueinrichtungen.
●
Kein Entkusseln (Birkenentfernen) um jeden Preis:
● Hydrologische Schutzzonen bzw. Klimaschutzzonen helfen Wasserstandsschwankungen auszugleichen sowie klimatische und Stickstoffbelastungen zu reduzieren (Pfadenhauer, Grootjans 1999; Edom 2001; Bönsel 2011).
● Randwälder helfen trophische und klimatische Einwirkungen zu reduzieren (Bretschneider 2012; Müller-Kröhling et al. 2019).
● Abschieben vorhandener Moorvegetation ist trotz Einsaat von Besenheide (Calluna vulgaris) erst bei Wassersättigung des Weißtorfkörpers zielführend.
5 Entwicklungen der Verlandungsstadien zur Hochmoorbultengesellschaft [2]
5.1 Zeiträume
Die Mehrzahl der Entwicklungen auf den Dauerflächen ist progressiv im Sinne des Leitbilds „Entwicklung einer Hochmoorbultengesellschaft“: Im Fockbeker, Wittenseer und Owschlager Moor sind bei unterschiedlichen trophischen Gegebenheiten und unterschiedlichstem hydrologischen
Management Ombrotrophierungstendenzen (Ernährung ausschließlich über atmosphärische Niederschläge) zu erkennen. Nach 10 – 30 Jahren haben sich alle der primären Wollgras- und Wasserschlauchstadien der Handtorfstiche zum Erico-Sphagnetum magellanici entwickelt (vgl.
Lütt 2001). Diese Ergebnisse stehen im Einklang mit 210Pb-Datierungen anhand von Torfprofilen verschiedener Torfstiche aus Mooren Schleswig-Holsteins (Lütt 1992) und Wiederholungskartierungen
(Frehse, Lütt 2005). Das entsprechende Zeitfenster für industriell abgetorfte Moorflächen ist ungleich größer (Schmatzler, Schmatzler 2010) und die Ausbreitung torfbildender Torfmoose schwieriger
(Lemmer, Graf 2016; Hölzel et al. 2019; Raabe et al. 2018).
5.2 Bedeutung der Stickstoffbelastung für Torfmoose
Die Ergebnisse zeigen, dass eine rezente Ausbreitung von Sphagnum magellanicum auf Kosten von S. fallax möglich ist (siehe Abb. 10). In den letzten 30 Jahren ist die Wirkung von Stickstoff auf
Sphagnen intensiv untersucht worden, ohne dass letztendlich klar daraus hervorgeht, warum die Ausbreitung von S. magellanicum trotz Konkurrenznachteilen bei erhöhten Stickstoffmengen noch erfolgt.

Abb. 10: Torfmoosrasen mit Sphagnum fallax, S. cuspidatum und S. magellanicum, das sich aktuell noch ausbreitet.
Fig. 10: Sphagnum lawns with S. fallax, S. cuspidatum and S. magellanicum, which is still expanding.
(Foto: Silke Lütt)
Aus Labor- und Felduntersuchungen ist bekannt, dass die Reaktion der Torfmoose auf ein höheres Stickstoffangebot unterhalb der Toxizitätsgrenze in einem gewissen Maße artspezifisch ist. Manche Arten, wie Sphagnum fallax (Twenhöven
1992; Risager 1998), S. lindbergii (Gunnarsson et al. 2004) oder ggf. auch S. papillosum (van der Heijden et al.
2000; vgl. auch Schmidt 2014) sind Konkurrenzstrategen, können selbst hohe Stickstoffangebote effektiv in Biomasse umsetzen. S. magellanicum hingegen scheint dieses Vermögen nicht oder nur begrenzt zu haben (Twenhöven 1992; Risager 1998). Heijmans et al. (2011) beobachtete wiederum bei Düngungsexperimenten mit 50 kg/ha–1 a–1 in
schwedischen Mooren an S. magellanicum im ersten Jahr zunächst Längenverluste aufgrund der Düngung, im dritten Jahr stellte sich allerdings infolge der Zunahme der Dichte eine höhere Produktivität ein. Die Dauer der Düngungsexperimente scheint die Wirkung ebenso zu
beeinflussen wie die Hintergrundbelastung, die am Standort der Versuchstorfmoose existierte und an die sie sich angepasst hatten (Bobbink, Hettelingh 2011). Darüber hinaus ist die Wirkung des Stickstoffs auch vom Mikrohabitat abhängig.
Torfmoosrasen und -teppiche reagieren weniger vulnerabel auf ein Überangebot an Stickstoff als Bulttorfmoose (Twenhöven 1992; Gunnarsson et al. 2000).
Im Kreis Rendsburg-Eckernförde, wo sämtliche Untersuchungsgebiete liegen, betragen die Hintergrundbelastungen zurzeit ca. 20 kg/ha–1 a–1 (Umweltbundesamt 2013 – 2015, siehe Schaap et al.
2018), d. h. sie betragen das Doppelte der Critical Loads (kritische Belastungsgrenzen) für Stickstoffeinträge in Mooren. Untersucht wurde in dieser Arbeit die Entwicklung von torfmoosdominierten Verlandungsstadien, die standörtlich weniger empfindlich auf
Stickstoffüberschüsse reagieren. Möglicherweise sind die kritischen Frachten mit toxischen Wirkungen bei diesen Belastungen noch nicht erreicht und es findet noch keine Co-Limitierung durch fehlenden Phosphor und fehlendes Kalium statt. Auch können Anpassungen von Sphagnum
magellanicum, z. B. durch höhere Dichten der Rasen, eine positive morphologische Anpassung an höhere Stickstoff-Angebote sein. Die wüchsigen Bestände des Wittenseer Moores mit hohen Deckungen von > 90 % an S. magellanicum und geringen
Deckungen an Gefäßpflanzen lassen vermuten, dass die Torfmoosrasen hier noch ihrer Filterfunktion gerecht werden und eine Immobilisierung des Stickstoffs ermöglichen.
5.3 Klimatische Effekte auf Torfmoose
Grundsätzlich wird davon ausgegangen, dass steigende Temperatur das Wachstum der Sphagnen stimuliert (z. B. Sonesson et al. 2002), wenn alle anderen Parameter im Optimum sind. Steigende Temperaturen bewirken allerdings zudem
sinkende Wasserstände und erhöhte Evapotranspiration, die dann umgekehrt eine Abnahme der Biomasseproduktion der Torfmoose (Gunnarsson et al. 2004; Breuuwer et al. 2009) und im Extremfall auch
schwere Trockenschäden bis zu einem Absterben der Sphagnen verursachen. Die bisherigen Temperaturerhöhungen von > 1 °C sind deshalb vermutlich noch unter der Schädigungsgrenze von Torfmoosen, zumal die Gesamtmenge der Niederschläge im Gebiet steigt. Allerdings gibt es
vermehrt Trockenphasen, insbesondere im Sommer.
Sphagnum magellanicum kann aufgrund seiner hohen Wasserhaltekapazität (Clymo, Hayword 1983) und höheren Austrocknungstoleranz im Vergleich zu S. fallax oder anderer Arten der Sektion Cuspidata (Dierßen, Dierßen
1984 in Ellenberg, Leuschner 2010) besser mit diesen veränderten Bedingungen umgehen. Stickstoffzufuhr erhöht die Dichte der Bulten zusätzlich (Heijmans et al. 2011). Gleichzeitig legt
S. magellanicum in Hinblick auf Massenzuwächse an den Bultstandorten im Vergleich zu den Cuspidaten zu (Lütt 1992) und gewinnt daher an Konkurrenzkraft. Abnehmende Wasserstände sind eine natürliche Folge des Aufwachsens der
Schwingdecken zu Bulten, also einer ungestörten Sukzession. Sie können aber auch durch das veränderte Klima beschleunigt werden. Die Ausbreitung von S. magellanicum auf vielen Dauerflächen könnte auch durch bessere Anpassungen an trockenere Standorte erfolgt sein.
5.4 Gesamtbetrachtung der Entwicklungen in allen drei Mooren
In allen drei Mooren konnten primäre Ombrotrophierungen nachgewiesen werden. Diese können durch steigende Temperaturen gefördert werden. Tahvanainen (2011) wies in finnischen Aapamooren eine Ombrotrophierung mit sich flächig ausbreitenden
Torfmoosen als Folge einer Entwässerung nach. Auch Schrautzer et al. (2019) konnten in österreichischen Mooren vergleichbare sekundäre Ombrotrophierungen belegen. In den untersuchten Mooren in Schleswig-Holstein gehen sie einher mit einer
deutlichen Zunahme der Deckung von Phanerogamen (Wittenseer Moor + 12 %, Fockbeker Moor + 30 %, Owschlager Moor + 48 %). Die Artenzahl ist in nahezu allen Dauerflächen gestiegen. Dies ist auf trockenere Standorte zurückzuführen, für Eriophorum
angustifolium, Vaccinium oxycoccos und Rhynchospora alba wurde jedoch auch eine Förderung durch Stickstoff nachgewiesen (Tüxen 1983; Heijmans et al. 2002; Wiedermann et al. 2007) und auch Narthecium ossifragum profitiert bei günstiger Kationenversorgung davon (Malmer et al. 2003).
Abnahmen der Torfmoosdeckung sind bei Schwingrasen zu verzeichnen, die stark an Phanerogamen und Laubmoosen zugenommen haben. Die Abnahme der Lebermoosarten (D6, Abb. 7; D8, Abb. 6,
Tab. B im Online-Zusatzmaterial II) und die Zunahme der Ericaceen, insbesondere von Calluna vulgaris, in allen Dauerflächen können ein
Hinweis auf die Verletzlichkeit der Hochmoorarten gegenüber dem Klimawandel sein (Sperle, Bruelheide 2020).
Eine Zunahme der Gefäßpflanzen stellten auch Gunnarsson et al. (2002) im Zeitraum von 1954 bis 1997 in einem Moor in Zentralschweden fest: einen Wandel hin zu einer etwas trockeneren und mehr an Stickstoff angepassten Vegetation mit
einer stärker ausgeprägten Mikrotopographie, vermehrten Bulttorfmoosen und beginnender Bewaldung.
6 Schlussfolgerungen
Die Ergebnisse bestätigen den Wert von Dauerbeobachtungen für Aussagen zum Erfolg von Renaturierungen. Das Ziel, hochmoortypische Vegetation wiederherzustellen, ist zurzeit noch möglich: wenn auch nicht auf den gesamten Moorflächen, so doch in Torfstichen und auf an- und
aufgestauten Teilbereichen.
Die festgestellte Akzentuierung der Mikrotopographie kann sowohl Spiegel einer autogenen Entwicklung als auch ein Hinweis auf erste Auswirkungen eines sich verändernden Klimas sein. Verstärkend wirken hohe Stickstofffrachten, die die Konkurrenz zwischen den Torfmoosen und
Gefäßpflanzen verändern und eine Schlenken- und Bultbildung vorantreiben. Der Konkurrenzvorteil von Sphagnum magellanicum, sich bei abnehmenden Wasserständen und zunehmenden Trockenheitsphasen aufgrund der hohen Dichte der Bulten behaupten zu können, kann seine durch Stickstoff
induzierte Konkurrenzschwäche möglicherweise ausgleichen.
Noch scheint die Grenze trophischer und klimatischer Belastungen für die Entstehung von Hochmoorbultengesellschaften nicht erreicht zu sein. Vor dem Hintergrund der bundes- wie landesweit seit Ende der 1990er-Jahre sinkenden Stickstoffoxidfrachten aus Verbrennungen der Industrie und
des Verkehrs (Schaap et al. 2018; Lorenz, Brunke 2021) besteht die Chance, dass dies auch in den nächsten Jahrzehnten nicht der Fall sein wird, wenn weitere naturschutzpolitische Vorgaben greifen.
Gleichwohl steigt angesichts der klimatischen Vulnerabilität der Hochmoorvegetation die Bedeutung eines adaptiven Managements selbst bei Prozessschutz (siehe Kasten 1). Insgesamt bestätigen die Ergebnisse den hohen Wert von
Renaturierungsmaßnahmen für die Wiederherstellung und Erhaltung einer spezifischen Hochmoorvegetation trotz hoher Stickstofffrachten und Klimaveränderungen.
Zusatzmaterial zum Beitrag
7 Literatur
↑
Aaby B. (1994): Monitoring Danish raised bogs. In: Grünig A. (Hrsg.): Mires and Man. Mire conservation in a densely populated country – The Swiss experience. Kosmos. Birmensdorf: 284 – 300.
↑
Belting S., Blankenburg J. (2014): Bericht zum Fachseminar „Perspektiven für den Moorschutz in Niedersachsen“. Telma 44: 171 – 180.
↑
Bobbink R., Hettelingh J.P. (Hrsg.) (2011): Review and revision of empirical critical loads and dose-response relationships. Proceedings of an expert workshop,
Noordwijkerhout, 23 – 25 June 2010. National Institute for Public Health and the Environment. Bilthoven: 243 S.
↑
Bönsel A. (2011): Revitalisierung von Regenmooren in Nordostdeutschland: Überblick und Perspektiven. Telma Beiheft 4: 27 – 48.
↑
Breeuwer A., Robroek B.J. et al. (2009): Decreased summer water table depth affects peatland vegetation. Basic and Applied Ecology 10(4):
330 – 339.
↑
Brehm K., Pfeifer G. (1982): Kann unseren Mooren noch geholfen werden? Rendsburger Jahrbuch 1982: 147 – 182.
↑
Bretschneider A. (2012): Die Bedeutung von Birken im Hochmoor. Telma 42: 137 – 146.
↑
Caspari S., Dürhammer O. et al. (2018): Rote Liste und Gesamtartenliste der Moose (Anthocerotophyta, Marchantiophyta und Bryophyta) Deutschlands. In: Metzing D., Hofbauer N. et al. (Red.): Rote Liste gefährdeter Tiere, Pflanzen und Pilze
Deutschlands. Band 7: Pflanzen. Naturschutz und Biologische Vielfalt 70(7): 361 – 489.
↑
Clymo R.S., Haword P.M. (1983): The Sphagnum ecology. In: Smith A.J. (Hrsg.): Bryophyte ecology. Chapman and Hall. London: 229 – 289.
↑
Daniels J. (2001): Ausbreitung der Moorbirke (Betula pubescens Ehrh. agg.) in gestörten Hochmooren der Diepholzer
Moorniederung. Osnabrücker Naturwissenschaftliche Mitteilungen 27: 39 – 49.
↑
Dierschke H. (1994): Pflanzensoziologie – Grundlagen und Methoden. Ulmer. Stuttgart: 683 S.
↑
Dierßen K., Glahn H. von et al. (1988): Rote Liste der Pflanzengesellschaften Schleswig-Holsteins. 2. Aufl. Schriftenreihe Landesamt für Naturschutz und Landschaftspflege Schleswig-Holstein 6: 157 S.
↑
Dierßen K., Nelle O. (2006): Zustand, Wandel und Entwicklung europäischer Moorlandschaften. Nova Acta Leopoldina NF 94(346): 241 – 257.
↑
DWD/Deutscher Wetterdienst (2017): Klimareport Schleswig-Holstein. Fakten bis zur Gegenwart – Erwartungen für die Zukunft. DWD. Offenbach a. M.: 40 S.
↑
Edom F. (2001): Revitalisierung von Regenmooren an ihrer klimatischen Arealgrenze. In: Succow M., Joosten H. (Hrsg.): Landschaftsökologische Moorkunde. E. Schweizerbart'sche Verlagsbuchhandlung. Stuttgart: 534 – 543.
↑
Ellenberg H., Leuschner C. (2010): Vegetation Mitteleuropas mit den Alpen. 6. Aufl. Ulmer. Stuttgart: 1 334 S.
↑
Finck P., Heinze S. et al. (2017): Rote Liste der gefährdeten Biotoptypen Deutschlands – dritte, fortgeschr. Fassung. Naturschutz und Biologische Vielfalt 156: 637 S.
↑
Frehse I., Lütt S. (2005): Vegetationsentwicklung im Nienwohlder Moor nach Restitutionsmaßnahmen. Mitteilungen der Arbeitsgemeinschaft Geobotanik in Schleswig-Holstein und Hamburg 65: 151 – 165.
↑
Gelbrecht J., Schiefelbein U. et al. (2011): Schutz der Moore in Mecklenburg-Vorpommern – Stand und Perspektiven. Telma Beiheft 4:
5 – 8.
↑
Gunnarsson U., Granberg G., Nilsson M. (2004): Growth, production and interspecific competition in Sphagnum: Effects of temperature, nitrogen and sulphur treatments on a boreal mire.
New Phytologist 163: 349 – 359.
↑
Gunnarsson U., Malmer N., Rydin H. (2002): Dynamics or constancy in Sphagnum dominated mire ecosystems? A 40-year study. Ecography 25:
685 – 704.
↑
Gunnarsson U., Rydin H., Sjörs H. (2000): Diversity and pH changes after 50 years on the boreal mire Skattlösbergs Stormosse, Central Sweden. Journal of Vegetation
Science 11(2): 277 – 286.
↑
Härdtle W., Assmann T. et al. (2009): Renaturierung und Management von Heiden. In: Zerbe S., Wiegleb G. (Hrsg.): Renaturierung von Ökosystemen in Mitteleuropa. Springer Spektrum. Heidelberg: 317 – 347.
↑
Heijmans M., Klees H. et al. (2002): Response of a Sphagnum bog plant community to elevated CO2 and N supply. Plant Ecology 162:
123 – 134.
↑
Heijmans M., Klees H., Berendse F. (2011): Competition between Sphagnum magellanicum and Eriophorum angustifolium as affected by raised
CO2 and increased N deposition. Oikos 97(3): 415 – 425.
↑
Heijmans M., van der Knaap Y.A. et al. (2013): Persistent versus transient tree encroachment of temperate peat bogs: Effects of climate warming and drought events. Global Change
Biology 19(7): 2 240 – 2 250.
↑
Hölzel, N., Kleinbecker T. et al. (2019): Leitfaden zur Torfmoosvermehrung für Renaturierungszwecke. Deutsche Bundesstiftung Umwelt. Osnabrück:
63 S.
↑
Kaule G., Carminati A. et al. (2018): Die Hochmoorwälder des süddeutschen Voralpengebietes: Moorökologische Bedeutung und zukünftige Entwicklung im Klimawandel. Telma 48:
13 – 48.
↑
Kaule G., Peringer A. (2015): Die Entwicklung der Übergangs- und Hochmoore im südbayrischen Voralpengebiet im Zeitraum 1969 – 2013 unter Berücksichtigung von Nutzungs- und Klimagradienten. Bayerisches Landesamt für Umwelt (LfU). Augsburg:
129 S.
↑
Kollmann J. (2019): Grundwasser- und Regenwassermoore. In: Kollmann J., Kirmer A. et al.: Renaturierungsökologie. Springer. Heidelberg: 171 – 192.
↑
Laube J. (2009): Die Revitalisierung der Moore im Steinwald. Ornithologischer Anzeiger 48: 36 – 42.
↑
Lemmer M., Graf M. (2016): Wie erfolgreich verläuft die Renaturierung abgebauter Hochmoore? Sphagnum-Vorkommen auf 19 wiedervernässten Flächen in Niedersachsen.
Telma 46: 109 – 124.
↑
LLUR/Landesamt für Landwirtschaft, Umwelt und ländliche Räume (Hrsg.) (2015): Moore in Schleswig-Holstein. Geschichte – Bedeutung – Schutz. Schriftenreihe LLUR SH – Natur 23: 161 S.
↑
Londo G. (1975): De decimale schaal foor vegetatiekundige opnamen van permanente Kwadraten. Gorteria 7(7): 101 – 106.
↑
Lorenz M., Brunke M. (2021): Trends of nutrients and metals in precipitation in northern Germany: The role of emissions and meteorology. Environmental Monitoring and
Assessment (2021): 325.
↑
Lütt S. (1992): Produktionsbiologische Untersuchungen zur Sukzession der Torfstichvegetation in Schleswig-Holstein. Mitteilungen der Arbeitsgemeinschaft Geobotanik in Schleswig-Holstein und Hamburg 43: 250 S.
↑
Lütt S. (2001): Die Veränderung der Vegetationsdecke von Torfstichen schleswig-holsteinischer Moore – Untersuchungen an Dauerbeobachtungsflächen. Kieler Notizen zur Pflanzenkunde in Schleswig-Holstein und Hamburg 29:
24 – 44.
↑
Lütt S. (2020): Vegetationsentwicklung in schleswig-holsteinischen Mooren in 30 Jahren. Langzeituntersuchungen an Dauerbeobachtungsflächen. Kieler Notizen zur Pflanzenkunde in Schleswig-Holstein und Hamburg 45:
7 – 50.
↑
Malmer N., Albinsson B. et al. (2003): Interferences between Sphagnum and vascular plants: Effects on plant community structure and peat
formation. Oikos 100(3): 469 – 482.
↑
Müller-Kröhling S., Schumacher J. et al. (2019): Beseitigung von Gehölzen im Moor. Naturschutz und
Landschaftsplanung 51(6): 264 – 269.
↑
Naturkapital Deutschland – TEEB DE (2014): Naturkapital und Klimapolitik – Synergien und Konflikte.
Kurzbericht für Entscheidungsträger. Technische Universität Berlin, Helmholtz-Zentrum für Umweltforschung – UFZ. Berlin, Leipzig: 216 S.
↑
Nick K.J., Weber H.E. (2001): Entwicklung der Vegetation auf dem wiedervernässten Leegmoor in den Jahren 1989 bis 1996. Angewandte Landschaftsökologie 38: 75 – 89.
↑
Pfadenhauer J, Grootjans A.P. (1999): Wetlands restoration in Central Europe: Aims and methods. Applied Vegetation Science 2(1): 95 – 106.
↑
Poschlod P. (1990): Vegetationsentwicklung in abgetorften Hochmooren des bayerischen Alpenvorlandes unter besonderer Berücksichtigung standortkundlicher und populationsbiologischer Faktoren. Dissertationes Botanicae 152: 331 S.
↑
Poschlod P. (1991): Diasporenbanken in Böden – Grundlagen und Bedeutung. Universität Hohenheim, Institut für Landeskultur und Pflanzenökologie. Stuttgart: 21 S.
↑
Raabe P., Kleinebecker T. et al. (2018): Vermehrung und Ansiedlung von Bulttorfmoosen in der Hochmoorrenaturierung – erste Ergebnisse eines Pilotprojekts im Landkreis
Vechta (Niedersachsen). Telma 48: 71 – 80.
↑
Rennwald E. (2000): Rote Liste der Pflanzengesellschaften Deutschlands. https://www.bfn.de/fileadmin/MDB/documents/RoteListePflanzengesellschaften.pdf (aufgerufen am 9.12.2020).
↑
Risager M. (1998): Impacts of nitrogen on Sphagnum dominated bogs with emphasis on critical load assessment. PhD thesis. University of Copenhagen. Copenhagen: 164 S.
↑
Sauer M., Ahrens M. (2006): Rote Liste und Artenverzeichnis der Moose Baden-Württembergs. Naturschutz-Praxis, Artenschutz 10. Landesanstalt für Umwelt, Messungen und Naturschutz Baden-Württemberg (LUBW). Karlsruhe: 142 S.
↑
Schaap M., Hendriks C. et al. (2018): PINETI-3: Modellierung atmosphärischer Stoffeinträge von 2000 bis 2015 zur Bewertung
der ökosystem-spezifischen Gefährdung von Biodiversität durch Luftschadstoffe in Deutschland. Abschlussbericht. Umweltbundesamt. Texte 79/2018. Dessau-Roßlau: 148 S.
↑
Schmatzler B., Schmatzler E. (2010): Moorland – Moorlandschaften in Niedersachsen nach industriellem Torfabbau. Bd. 1. Industrieverband Garten e. V. Ratingen: 465 S.
↑
Schmidt S.R. (2014): Sphagnum in a changing world: From the landscape to the isotope scale. Dissertation Universität Hamburg. Hamburg:
141 S.
↑
Schrautzer J., Bork H.R. et al. (2019): Classification,
ecological characterization and development of montane mires (Kleinwalsertal, Austria). Phytocoenologia 49(3): 263 – 285.
↑
Sliva J. (1997): Renaturierung von industriell abgetorften Hochmooren am Beispiel des Kendlmühlfilzen. Herbert Utz Verlag. München: 229 S.
↑
Sonesson M., Carlsson B.A. et al. (2002): Growth of two peat-forming mosses in subarctic mires: Species interactions and effects of simulated
climate change. Oikos 99(1): 151 – 160.
↑
Sperle T., Bruelheide H. (2020): Climate change aggravates bog species extinctions in the Black Forest (Germany). Diversity and Distributions 27(2):
282 – 295.
↑
Ssymank A., Hauke U. et al. (1998): Das europäische Schutzgebietssystem NATURA 2000. Schriftenreihe für Landschaftspflege und Naturschutz 53: 560 S.
↑
Ssymank A., Ullrich K. et al. (2015): Handlungsleitfaden Moorschutz und Natura 2000 für die Durchführung von Moorrevitalisierungsprojekten. In: Vischer-Leopold M., Ellwanger G. et al. (2015): Natura 2000 und Management in
Moorgebieten. Naturschutz und Biologische Vielfalt 140: 277 – 312.
↑
Succow M., Joosten H. (2001): Landschaftsökologische Moorkunde. E. Schweizerbart'sche Verlagsbuchhandlung. Stuttgart: 622 S.
↑
Tahvanainen T. (2011): Abrupt ombrotrophication of a boreal aapa mire triggered by hydrological disturbance in the catchment. Journal of
Ecology 99(2): 404 – 415.
↑
Timmermann T., Joosten H., Succow M. (2009): Restaurierung von Mooren. In: Zerbe S., Wiegleb G. (Hrsg.): Renaturierung von Ökosystemen in Mitteleuropa. Springer Spektrum. Heidelberg: 55 – 93.
↑
Tüxen J. (1983): Die Schutzwürdigkeit der niedersächsischen Kleinstmoore im Hinblick auf ihre Vegetation. Tüxenia 3:
423 – 435.
↑
Twenhöven F. (1992): Untersuchungen zur Wirkung stickstoffhaltiger Niederschläge auf die Vegetation von Hochmooren. Mitteilungen der Arbeitsgemeinschaft Geobotanik in Schleswig-Holstein und Hamburg 44: 172 S.
↑
Van Breemen N. (1995): How Sphagnum bogs down other plants. Trends in Ecology and Evolution 10(7): 270 – 275.
↑
Van der Heijden E., Jauhiainen J. et al. (2000): Effects of elevated atmospheric CO2 concentration and increased nitrogen deposition on growth and
chemical composition of ombrotrophic Sphagnum balticum and oligo-mesotrophic Sphagnum papillosum. Journal of Bryology 22(3): 175 – 182.
↑
Waesch G. (2006): Untersuchungen zum Diasporenvorrat unterschiedlich genutzter Wiesen im Thüringer Wald – kann die Samenbank Regeneration
von Grasland bewirken? Tuexenia 26: 275 – 295.
↑
Wagner C. (1994): Zur Ökologie der Moorbirke Betula pubescens Ehrh. in Hochmooren Schleswig-Holsteins unter besonderer Berücksichtigung von Regenerationsprozessen in Torfstichen. Mitteilungen der Arbeitsgemeinschaft
Geobotanik in Schleswig-Holstein und Hamburg 47: 182 S.
↑
Wiedermann M.M., Nordin A. et al. (2007): Global change shifts vegetation and plant-parasite interactions in a boreal mire. Ecology 88(2):
454 – 464.
↑
Wisskirchen R., Haeupler H. (1998): Standardliste der Farn- und Blütenpflanzen Deutschlands. Ulmer. Stuttgart: 765 S.
Fußnoten
↑
[1] Wesentliche Teile des Kapitels entsprechen weitgehend Lütt (2020).
↑
[2] Wesentliche Teile des Kapitels entsprechen weitgehend Lütt (2020).

